Médico especialista do artigo

Novas publicações
Síntese, secreção e metabolismo de catecolaminas
Última revisão: 19.10.2021

Todo o conteúdo do iLive é medicamente revisado ou verificado pelos fatos para garantir o máximo de precisão factual possível.
Temos diretrizes rigorosas de fornecimento e vinculamos apenas sites de mídia respeitáveis, instituições de pesquisa acadêmica e, sempre que possível, estudos médicos revisados por pares. Observe que os números entre parênteses ([1], [2], etc.) são links clicáveis para esses estudos.
Se você achar que algum dos nossos conteúdos é impreciso, desatualizado ou questionável, selecione-o e pressione Ctrl + Enter.
A camada cerebral das glândulas supra-renais produz um composto longe dos esteróides da estrutura. Eles contêm um núcleo de 3,4-di-hidroxifenil (catecol) e são chamados catecolaminas. Estes incluem adrenalina, norepinefrina e dopamina beta-oxitiramina.
Síntese da catecolamina sequência é bastante simples: Tirosina → di-hidroxifenilalanina (DOPA) → → dopamina norepinefrina → adrenalina. A tirosina entra no organismo com alimentos, mas também pode ser formada a partir da fenilalanina no fígado, sob a ação da fenilalanina hidroxilase. Os produtos finais da transformação da tirosina nos tecidos são diferentes. Na medula supra-renal o processo prossegue para o passo de formação de adrenalina, nas extremidades dos nervos simpáticos - noradrenalina, em certos neurónios do sistema nervoso catecolaminas síntese de dopamina formulário preenchido central.
A conversão da tirosina em DOPA é catalisada pela tirosina hidroxilase, cofatores dos quais são tetrahidro-biopterina e oxigénio. Acredita-se que é essa enzima que limita a velocidade de todo o processo de biossíntese de catecolaminas e é inibida pelos produtos finais do processo. A tirosina hidroxilase é o principal objeto de efeitos regulatórios na biossíntese de catecolaminas.
Dopa para conversão dopamina é catalisada pela enzima de dopa-descarboxilase (cofactor - piridoxal), que é relativamente não específico e descarboxilado, e outra aromático L-amino ácido. No entanto, existem indícios da possibilidade de modificar a síntese de catecolaminas, alterando a atividade e esta enzima. Em alguns neurônios não há enzimas para a conversão adicional da dopamina, e é o produto final. Outros tecidos contêm dopamina-beta-hidroxilase (cofatores são cobre, ácido ascórbico e oxigênio), que converte a dopamina em noradrenalina. Na medula supra-renal (mas não terminações nervosas simpáticas) é feniletanolamina presente - formando adrenalina de noradrenalina metiltransferase. O dador dos grupos metilo neste caso é S-adenosilmetionina.
É importante lembrar que a síntese de feniletanolamina-N-Metiltransferazy induzida por glucocorticóides que caem na camada cortical cerebral do sistema venoso portal. Isto pode explicar as mentiras fato combinando duas glândulas endócrinas diferentes em um só corpo. Significado da síntese de adrenalina glucocorticóide enfatizada pelo facto das células de medula adrenal produzir norepinefrina, dispostos em torno dos vasos arteriais, enquanto que as células do sangue são obtidos adrenalinprodutsiruyuschie essencialmente de seios venosos, localizada no córtex supra-renal.
O colapso de catecolaminas ocorre principalmente sob a influência de dois sistemas de enzimas: catecol-O-metiltransferase (COMT) e monoamina-oxidase (MAO). As principais maneiras de epinefrina e norepinefrina de decaimento mostrado esquematicamente na Fig. 54. Sob a acção da COMT na presença de um dador de catecolaminas grupos metilo S-adrenozilmetionina e convertido em metanefrina normetanefrina (3-O-metil-derivados de epinefrina e norepinefrina), que sob a influência da MAO transformados em aldeídos e mais (na presença de aldeído) em vanilil-mandélico ácido (ICH) - o produto de degradação principal de noradrenalina e adrenalina. No mesmo processo, quando pela primeira vez exposto à acção da MAO catecolaminas, não COMT, que são convertidos para o aldeído 3,4-dioksimindalevy, e, em seguida, sob a influência do aldeído e da COMT - ácido 3,4-dioksimindalnuyu e o DIU. Na presença de álcool-desidrogenase de catecolaminas podem formar-3-metoxi-4 oksifenilglikol, o principal produto final da degradao de epinefrina e norepinefrina no SNC.
Desintegração dopamina prossegue de modo semelhante, excepto que os seus metabolitos são desprovidos de grupos hidroxilo no átomo de carbono beta e, por isso, em vez de ácido vanílico-mandélico homovanílico formado (AHV) e ácido 3-metoxi-4-oksifeniluksusnaya.
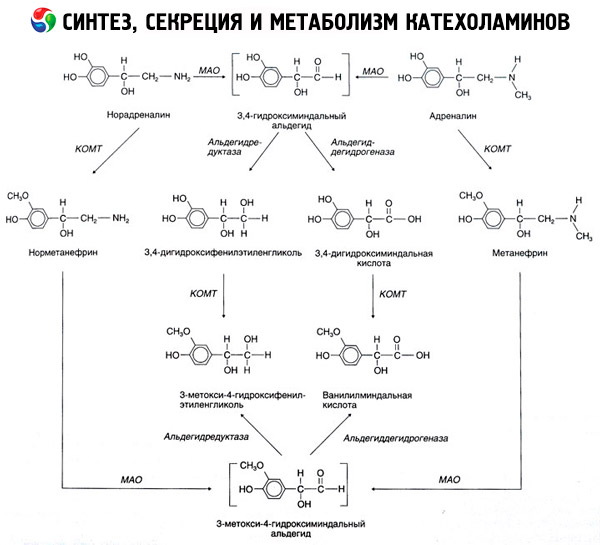
A existência de uma via quinoide para a oxidação da molécula de catecolaminas, na qual também podem ser postulados produtos intermediários com uma atividade biológica pronunciada.
Formada pela acção de enzimas citosólicas, adrenalina e noradrenalina nas terminações nervosas simpáticas, medula adrenal e introduzir os grânulos de secreção, que as protege contra a acção de enzimas degradativas. A captura de catecolaminas com grânulos requer custos energéticos. Em grânulos cromafins das catecolaminas medula adrenal firmemente ligados ao ATP (na proporção de 4: 1) e as proteínas específicas - cromogranina que evita a difusão de hormonas a partir dos grânulos no citoplasma.
Estímulo directo à secreção de catecolaminas é aparentemente a penetração celular cálcio estimular exocitose (grânulos de fusão de membrana com a superfície da célula e a sua abertura com o rendimento global do conteúdo solúvel - catecolaminas, dopamina-beta-hidroxilase, ATP e cromogranina - para o fluido extracelular) .
Efeitos fisiológicos das catecolaminas e o mecanismo de sua ação
Os efeitos das catecolaminas começam com a interação com receptores específicos de células alvo. Se os receptores de hormônios tireoideanos e esteróides estiverem localizados dentro das células, os receptores de catecolaminas (assim como os hormônios acetilcolina e peptídeo) estão presentes na superfície celular externa.
Tem sido demonstrado que, em relação a algumas reacções adrenalina ou noradrenalina são mais eficazes do que o isoproterenol catecolamina sintética, enquanto que para outros, o efeito é superior à acção de epinefrina ou isoproterenol norepinefrina. Com base neste conceito para a presença de dois tipos de tecidos foi desenvolvido adrenérgicos alfa e beta, e apenas qualquer um destes dois tipos pode estar presente em alguns deles. Isoproterenol é o agonista mais potente dos receptores beta-adrenérgicos, enquanto que o composto fenilefrina sintético - o agonista mais potente dos receptores alfa-adrenérgicos. Catecolaminas naturais - adrenalina e noradrenalina - são capazes de interagir com os receptores de ambos os tipos, mas a adrenalina toma uma maior afinidade para o beta, e norepinefrina - alfa-receptores.
Catecolaminas fortes activar os receptores beta-adrenérgicos cardíacos do que os receptores beta do músculo liso, o que permite a-tipo beta divididos em subtipos: receptores beta1 (coração, células de gordura) e receptores beta-2 (brônquios, vasos sanguíneos, etc ...). A acção de isoproterenol no receptor beta 1 de acção superior de adrenalina e noradrenalina apenas 10 vezes, enquanto que os receptores beta-2 que actua 100-1000 vezes mais potente do que a catecolamina natural.
Os antagonistas específicos de aplicação (fenoxibenzamina e fentolamina contra alfa e beta propranolol contra-receptor) confirmaram a adequação da classificação dos receptores adrenérgicos. A dopamina é capaz de interagir tanto com o alfa e beta-receptores, mas em vários tecidos (cérebro, glândula pituitária, vasos) encontrado e receptores dopaminérgicos próprios bloqueador específico que é o haloperidol. O número de receptores beta varia de 1000 a 2000 por célula. Efeitos biológicos das catecolaminas mediadas por beta-receptores estão ligados, por via de regra, com a activação da adenilato ciclase e cAMP aumentado intracelular. O receptor e a enzima, embora estejam conectados funcionalmente, mas representam diferentes macromoléculas. A modulação da actividade da adenilato-ciclase, sob a influência do complexo hormona-receptor são trifosfato de guanosina envolvido (GTP) e outros nucleótidos de purina. Ao aumentar a atividade da enzima, eles parecem reduzir a afinidade dos receptores beta por agonistas.
O fenômeno de aumentar a sensibilidade de estruturas desnervadas é conhecido há muito tempo. Por outro lado, a exposição prolongada a agonistas diminui a sensibilidade dos tecidos-alvo. O estudo dos receptores beta permitiu explicar esses fenômenos. Tem sido demonstrado que a exposição prolongada ao isoproterenol provoca uma perda de sensibilidade da adenilato ciclase através da redução do número de beta-receptores.
O processo de dessensibilização não requer a ativação da síntese protéica e provavelmente se deve à formação gradual de complexos receptores hormonais irreversíveis. Pelo contrário, a administração da 6-oxidofamina, que quebra as extremidades simpáticas, é acompanhada por um aumento no número de receptores beta reagentes nos tecidos. Não está excluído que um aumento na atividade nervosa simpática determine a dessensibilização relacionada à idade dos vasos sanguíneos e do tecido adiposo em relação às catecolaminas.
O número de adrenorreceptores em diferentes órgãos pode ser controlado por outros hormônios. Assim, ela aumenta o estradiol, progesterona e reduzem o número de receptores alfa-adrenérgicos no útero, que é acompanhado por um aumento e diminuição da sua resposta contráctil à catecolaminas correspondente. Se intracelular "segundo mensageiro", formados pela acção dos agonistas dos receptores beta, certamente é AMPc, em relação ao do transmissor de efeitos alfa-adrenérgicos é mais complicado. A existência de vários mecanismos: a redução dos níveis de cAMP, aumento dos níveis de cAMP, modulação da dinâmica celular de cálcio, e outros.
Para reproduzir uma variedade de efeitos no corpo, doses de epinefrina, que são 5-10 vezes menores que a norepinefrina, são geralmente necessárias. Embora o último é mais eficaz no que diz respeito ao a- e os receptores beta1-adrenérgicos, é importante lembrar que ambas catecolamina endógena capaz de interagir tanto com o alfa e beta-receptores. Portanto, a resposta biológica desse corpo à ativação adrenérgica depende em grande parte do tipo de receptores presentes nele. No entanto, isso não significa que a ativação seletiva do elo nervoso ou humoral do sistema simpático-adrenal seja impossível. Na maioria dos casos, há uma atividade intensificada de seus vários elos. Assim, presume-se que ele ativa hipoglicemia reflexo medula supra-renal, ao passo que uma diminuição da pressão arterial (hipotensão postural), acompanhada de libertação principalmente norepinefrina a partir dos terminais dos nervos simpáticos.
Adrenorreceptores e os efeitos de sua ativação em vários tecidos
|
Sistema, órgão |
Tipo de adrenorreceptor |
Reação |
|
Sistema cardiovascular: |
||
|
Coração |
Beta |
Aumento na freqüência de contrações, condução e contratilidade |
|
Arteríolas: |
||
|
Pele e membranas mucosas |
Alfa |
Redução |
|
Dos músculos esqueléticos |
Beta |
Redução de Extensão |
|
órgãos abdominais |
Alfa (mais) |
Redução |
|
Beta |
Extensão |
|
|
Veias |
Alfa |
Redução |
|
Sistema respiratório: |
||
|
Músculos bronquiais |
Beta |
Extensão |
|
O sistema digestivo |
||
|
Estômago |
Beta |
Função motora diminuída |
|
Intestino |
Alfa |
Redução de esfíncteres |
|
Baço |
Alfa |
Redução |
|
Beta |
Relaxamento |
|
|
Parte externamente secreta do pâncreas |
Alfa |
Secreção diminuída |
|
Sistema geniturinário: |
Alfa |
Redução do esfíncter |
|
Bexiga |
Beta |
Relaxando o músculo exorcista |
|
Órgãos sexuais masculinos |
Alfa |
Ejaculação |
|
Olhos |
Alfa |
Pupila dilatada |
|
Couro |
Alfa |
Sudorese aumentada |
|
Glândulas salivares |
Alfa |
Isolamento de potássio e água |
|
Beta |
Secreção de amilase |
|
|
Glândulas endócrinas: |
||
|
Ilhotas do pâncreas |
||
|
Células beta |
Alfa (mais) |
Secreção de insulina diminuída |
|
Beta |
Aumento da secreção de insulina |
|
|
Células alfa |
Beta |
Aumento da secreção de glucagon |
|
8 células |
Beta |
Aumento da secreção de somatostatina |
|
O hipotálamo e a pituitária: |
||
|
Somatotróficos |
Alfa |
Aumento da secreção de STH |
|
Beta |
Redução da secreção de STH |
|
|
Lactotrofos |
Alfa |
Diminuição da secreção de prolactina |
|
Tirotrofos |
Alfa |
Diminuição da secreção de TSH |
|
Corticotrofos |
Alfa |
Aumento da secreção de ACTH |
| beta | Secreção diminuída de ACTH | |
|
Glândula tireóide: |
||
|
Células foliculares |
Alfa |
Diminuição da secreção de tiroxina |
|
Beta |
Aumento da secreção de tiroxina |
|
|
Células parafoliculares (K) |
Beta |
Aumento da secreção de calcitonina |
|
Glândulas paratireoides |
Beta |
Aumento da secreção de PTH |
|
Rins |
Beta |
Aumento da secreção de renina |
|
Estômago |
Beta |
Aumentar a secreção de gastrina |
|
Troca Básica |
Beta |
Aumento no consumo de oxigênio |
|
O fígado |
? |
Aumento na glicogenólise e na gliconeogênese a partir do rendimento de glicose; aumentar a cetogênese com a liberação de corpos cetônicos |
|
Tecido adiposo |
Beta |
O aumento da lipólise com a liberação de ácidos graxos livres e glicerol |
|
Músculos esqueléticos |
Beta |
Aumento da glicólise com a liberação de piruvato e lactato; diminuição na proteólise com uma diminuição no rendimento de alanina, glutamina |
É importante ter em mente que os resultados da administração intravenosa de catecolaminas não são sempre refletem adequadamente os efeitos de compostos endógenos. Isso se aplica principalmente a noradrenalina, porque ele está na sua maioria não o corpo do sangue, mas diretamente na fenda sináptica. Portanto norepinefrina endógena é activado, por exemplo, não só os receptores alfa vasculares (aumento da pressão sanguínea), mas também de beta-receptores cardíacos (palpitação), enquanto que a administração de fios externos noradrenalina predominantemente para a activação do receptor alfa vascular e reflexo (via o nervo vago) abrandamento batimentos cardíacos.
Baixas doses de epinefrina ativam principalmente os receptores beta dos vasos musculares e cardíacos, resultando em queda da resistência vascular periférica e aumento do volume minuto do coração. Em alguns casos, o primeiro efeito pode predominar e, após a administração de adrenalina, desenvolve-se hipotensão. Em doses mais altas, a adrenalina também ativa os receptores alfa, que é acompanhada por um aumento na resistência vascular periférica e, no contexto de um aumento no volume diminuto do coração, leva a um aumento na pressão sanguínea. No entanto, seu efeito sobre os receptores beta vasculares também é preservado. Como resultado, o aumento da pressão sistólica excede o valor similar da pressão diastólica (aumento da pressão de pulso). Com a introdução de doses ainda maiores, os efeitos alfa-miméticos da epinefrina começam a prevalecer: o aumento da pressão sistólica e diastólica em paralelo, ambos sob a influência da noradrenalina.
O efeito das catecolaminas no metabolismo é constituído pelos seus efeitos diretos e indiretos. Os primeiros são realizados principalmente através de receptores beta. Processos mais complexos estão associados ao fígado. Embora o aumento da glicogenólise hepática tenha sido tradicionalmente considerado o resultado da ativação do receptor beta, também há dados sobre o envolvimento dos receptores alfa neste. Os efeitos mediados das catecolaminas estão associados à modulação da secreção de muitos outros hormônios, por exemplo, a insulina. Na ação da adrenalina em sua secreção, o componente alfa-adrenérgico predomina claramente, pois é demonstrado que qualquer estresse é acompanhado pela inibição da secreção de insulina.
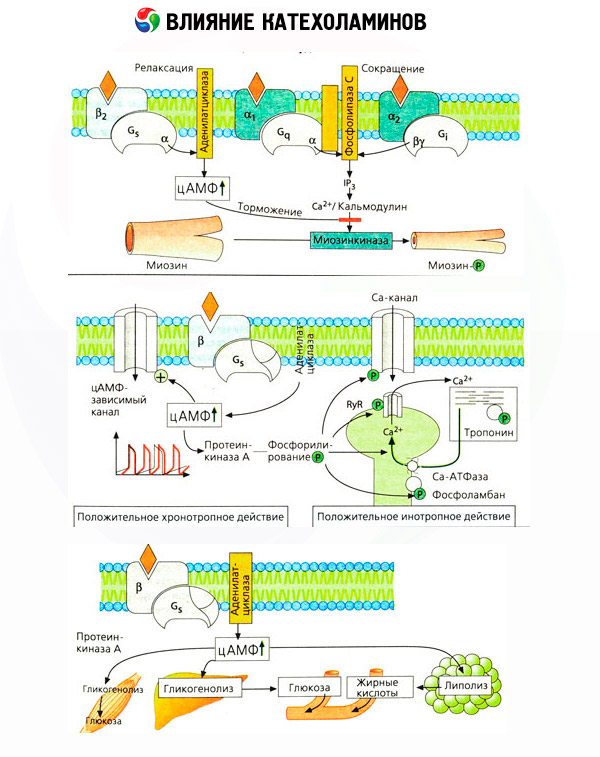
A combinação de efeitos diretos e indiretos das catecolaminas causa hiperglicemia, associada não somente ao aumento da produção hepática de glicose, mas também à inibição de sua utilização pelos tecidos periféricos. A aceleração da lipólise causa hiperlipacidemia com aumento da oferta de ácidos graxos ao fígado e intensificação da produção de corpos cetônicos. O aumento da glicólise nos músculos leva a um aumento na liberação de lactato e piruvato no sangue, que juntamente com o glicerol liberado do tecido adiposo servem como precursores da gliconeogênese hepática.
Regulação da secreção de catecolaminas. A similaridade dos produtos e métodos de resposta do sistema nervoso simpático e a medula supra-renal foi a base para a combinação destas estruturas em um único corpo simpatoadrenal neural libertação sistema hormonal e a sua ligação. Vários sinais aferentes estão concentrados no hipotálamo e centros da medula e da medula oblongata vertebral a partir da qual emanam comutação parcela eferente sobre corpos celulares neurónios pré-ganglionares localizados no corno lateral da medula espinal ao nível da VIII cervical - segmentos lombares II-III.
Axónios pré-ganglionares destas células estão deixando a medula espinhal e formar conexões sinápticas com os neurónios são localizadas nos gânglios da cadeia simpática, ou as células de medula adrenal. Estes são fibras pré-ganglionares colinérgicos. A primeira diferença fundamental de neurónios simpáticos e medula adrenal pós-ganglionares células cromafins consiste em que este último é transmitido o sinal de chegada que neuro-condução colinérgica (nervos adrenérgicos pós-ganglionares) e humoral, destacando composto adrenérgico para o sangue. A segunda diferença é reduzida para nervos pós-ganglionares que produzem norepinefrina, ao passo que as células de medula adrenal - de preferência adrenalina. Estas duas substâncias têm efeitos diferentes sobre o tecido.