Médico especialista do artigo

Novas publicações
Ovário
Última revisão: 23.04.2024

Todo o conteúdo do iLive é medicamente revisado ou verificado pelos fatos para garantir o máximo de precisão factual possível.
Temos diretrizes rigorosas de fornecimento e vinculamos apenas sites de mídia respeitáveis, instituições de pesquisa acadêmica e, sempre que possível, estudos médicos revisados por pares. Observe que os números entre parênteses ([1], [2], etc.) são links clicáveis para esses estudos.
Se você achar que algum dos nossos conteúdos é impreciso, desatualizado ou questionável, selecione-o e pressione Ctrl + Enter.
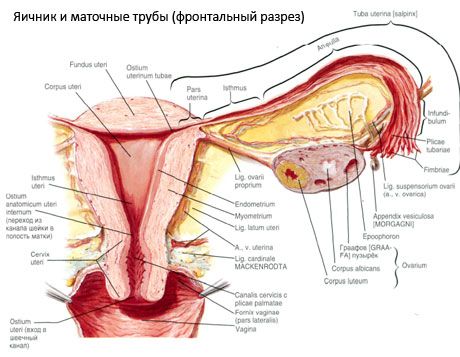
Ovário (ovário, oophoron grego) - órgão emparelhado, glândula genital feminina, localizado na cavidade da pequena pelve atrás do ligamento largo do útero. Os ovários desenvolvem células sexuais femininas maduras (óvulos), bem como hormônios sexuais femininos que entram no sangue e na linfa. O ovário tem uma forma ovoide, um tanto achatada na direção anterior-posterior. A cor do ovário é rosada. Na superfície do ovário de uma mulher que dá nascimento, depressões e cicatrizes são visíveis - traços de ovulação e transformação de corpos amarelos. A massa do ovário é de 5-8 g. As dimensões do ovário são 2,5-5,5 cm de comprimento, 1,5-3,0 cm de largura e 2 cm de espessura. O ovário tem duas superfícies livres: a facies medialis ), de frente para a cavidade da pequena pélvis, parcialmente coberta pela trompa de Falópio, e a superfície lateral (facies lateralis), adjacente à parede lateral da pequena pelve, à depressão ligeiramente expressa - a fossa ovariana. Esta fóvea fica na esquina entre os vasos ilíacos externos peritoneais na parte superior e as artérias uterinas e oclusais na parte inferior. Atrás do ovário, o ureter do lado correspondente passa da parte superior para a parte inferior.
Superfície do ovário em movimento numa livre (traseira) borda (liber margo) convexa, em frente - na região mesentérica (margo mesovaricus), por meio de um grampo curto-dobra do peritoneu (ovário mesentério) para a folha de trás do ligamento largo do útero. Nesta extremidade anterior do órgão há uma depressão ranhurada - o portão do ovário (hilum ovarii), através do qual a artéria e os nervos entram no ovário, as veias e os vasos linfáticos saem. O ovário também tem duas extremidades: uma extremidade do tubo superior arredondada (extrema tubaria) voltada para a trompa de Falópio e uma extremidade uterina inferior (extremitas utenna) conectada ao útero por um ligamento do ovário (Ovarii proprium). Este feixe na forma de uma cadeia circular de cerca de 6 mm de espessura vai da extremidade uterina do ovário ao canto lateral do útero, localizado entre duas folhas do ligamento amplo do útero. O aparelho ligamentoso do ovário também inclui um ligamento suspenso do ovário (lig.suspensorium ovarii), que é uma dobra do peritônio que se desloca de cima da parede pélvica para o ovário e que contém dentro dos vasos ovarianos e feixes de fibras fibrosas. O ovário é fixado com um mesentério curto (mesovário), que é uma duplicação peritoneal que corre da folha posterior do ligamento amplo do útero para a margem mesentérica do ovário. Os próprios ovários não são cobertos pelo peritoneu. A maior fimbria do ovário do tubo uterino é anexada à extremidade do tubo do ovário. A topografia do ovário depende da posição do útero, sua magnitude (durante a gravidez). Os ovários se referem a órgãos muito móveis da cavidade pélvica.
 [
[Vasos ovarianos e nervos
O suprimento de sangue para os ovários é devido à aa. Et vv. Ovaricae e uterinae. Ambas as artérias ováricas (aa. Ovaricae dextra et sinistra) partem da superfície anterior da aorta logo abaixo das artérias renais, a direita mais frequentemente proveniente da aorta e da esquerda da artéria renal. Indo para baixo e lateralmente sobre o músculo psoas superfície frontal, artéria ovário intercepta ureter cada frente (dando-lhe galhos), os vasos ilíacos externos, e a linha de fronteira vêm para dentro da cavidade pélvica, sendo localizado aqui no ligamento suspensor do ovário. Seguindo a direção medial, a artéria ovariana passa entre as folhas do ligamento amplo do útero sob a trompa de Falópio, dando-lhe ramos, e depois para o mesentério do ovário; entra nos portões do ovário.
Os ramos da artéria ovariana são amplamente anastomosados com os ramos ovarianos da artéria uterina. A saída venosa dos ovários é realizada principalmente no plexo venoso ovariano, localizado na área dos portões dos ovários. Daí a saída de sangue passa em duas direções: através das veias uterinas e ovarianas. A veia ovariana direita possui válvulas e corre para a veia cava inferior. A veia ovariana esquerda flui para a veia renal esquerda, sem válvulas presentes.
A saída linfática dos ovários ocorre através dos vasos linfáticos, especialmente abundantes na área dos portões dos órgãos, onde o plexo linfático sublingual é excretado. Então, a linfa é desviada para os linfonodos para-aórticos ao longo dos vasos linfáticos ovarianos.
Inervação dos ovários
Simpático - é fornecido por fibras pós-ganglionares dos plexos celíacos (solares), trinados superiores e hipogástricos; parasimpático - devido aos nervos sacrais internos.
Estrutura do ovário
A superfície do ovário é coberta com um epitélio germinal de camada única. Por baixo existe um envelope denso de tecido conjuntivo (túnica albugínea). O tecido conjuntivo do ovário forma seu estroma (stroma ovarii), rico em fibras elásticas. A substância do ovário, seu parênquima, é dividida nas camadas externa e interna. A camada interna situada no centro do ovário, mais perto de seus portões, é chamada medula ovarii. Nesta camada no tecido conjuntivo solto existem numerosos vasos sanguíneos e linfáticos e nervos. A camada externa do ovário do córtex do ovário é mais densa. Tem uma grande quantidade de tecido conjuntivo em que estão localizados de maturação de folículos do ovário primárias (folliculi ovarici primarii), folículos (bolha) secundário (folliculi ovarici secundarii, s.vesiculosi), um bem folículos maduros graafovy bolhas (folliculi ovarici maturis), e amarelo e corpos atretic.
Em cada folículo é um óvulo reprodutivo feminino, ou oócito (ovócito). Ovário com um diâmetro de até 150 μm, arredondado, contém um núcleo, uma grande quantidade de citoplasma, em que, além das organelas celulares, existem inclusões de proteína-lípido (gema), glicogênio necessário para alimentar o ovo. O suprimento de oócitos geralmente consome dentro de 12-24 horas após a ovulação. Se a fertilização não ocorrer, o óvulo morre.
O ovário humano tem duas membranas de cobertura. O interior é o citomema, que é a membrana citoplasmática do oócito. Fora do citomema, existe uma camada das chamadas células foliculares que protegem o ovo e cumprem a função formadora de hormônio - liberação de estrogênio.
A posição fisiológica do útero, dos tubos e dos ovários é fornecida pelo aparelho de suspensão, fixação e suporte, combinando peritoneu, ligamentos e celulose pélvica. O dispositivo de suspensão é representado por formações emparelhadas, inclui ligamentos redondos e largos do útero, ligamentos próprios e ovários ligamentares pendurados. O ligamento largo do útero, o ligamento dos ovários e de suspensão mantém o útero na posição intermediária. Os ligamentos redondos atraem a parte inferior do útero anteriormente e proporcionam sua inclinação fisiológica.
O dispositivo de fixação assegura a posição do instável no centro da pequena pélvis e torna praticamente impossível deslocá-la para os lados, para frente e para trás. Mas como o aparelho ligamentoso se afasta do útero na parte inferior, é possível inclinar o útero em várias direções. Pela unidade de fixação inclui cabos dispostos no tecido pélvico solto e que se estendem desde a placa inferior do útero para o lado, paredes dianteira e traseira da pelve: sacroiliac magochnye cardinal, uterino e ligamento cística-vesico púbico.
Além do mesovarium, distinguem-se os seguintes ligamentos ovarianos:
- uma suspensão do ligamento do ovário, anteriormente referida como voronkotazovaya. É uma dobra do peritoneu com que se prolonga nela sangue (um. Et v. Ovárica) e vasos linfáticos e nervos de ovário de esticado entre a parede pélvica lateral, fáscia lombar (em dividindo a artéria ilíaca comum no exterior e interior) e superior (tubo) o fim do ovário;
- o ligamento do ovário passa entre os lenços do ligamento uterino largo, mais próximo da folha posterior e conecta o extremo inferior do ovário com o margem lateral do útero. Para o útero, o ligamento do ovário está ligado entre o início do tubo uterino e o ligamento circular, para trás e para trás a partir deste último. Na espessura do ligamento são rr. Ovarii, que são os ramos terminais da artéria uterina;
- Ligamento apendicular-ovariano O clado se estende sobre o cume do apêndice para o ovário direito ou o ligamento largo do útero na forma de uma dobra do peritoneu. O ligamento é instável e é observado em 1/2 - 1/3 de mulheres.
O aparelho de suporte é representado pelos músculos e fascias do assoalho pélvico, divididos em camadas inferior, média e superior (interna).
O mais poderoso é a camada muscular superior (interna), representada pelo músculo pareado que eleva o ânus. Consiste em feixes de músculos que se afastam do cóccix para os ossos da pelve em três direções (músculos pubic-coccygeal, ilio-coccygeal e isquiococcígeo). Esta camada de músculos também é chamada de diafragma da pelve.
A camada intermediária dos músculos está localizada entre a sínfise, os ossos do púbis e do isquio. A camada média dos músculos - o diafragma urogenital - ocupa a metade anterior da saída pélvica, através da passagem da uretra e da vagina. Na parte anterior entre as suas folhas estão os feixes musculares que formam o esfíncter externo da uretra, na parte traseira há feixes musculares indo na direção transversal, o músculo transversal profundo do periné.
A camada inferior (externa) dos músculos do assoalho pélvico consiste em músculos da superfície, cuja forma se assemelha à figura 8. Estes incluem bulbo-cavernoso, isquial-cavernoso, esfíncter externo do ânus, músculo perineal transversal superficial.
Ontogenia dos ovários
O processo de crescimento e a atresia folicular começa com 20 semanas de gravidez e, no momento do parto nos ovários, a menina permanece até 2 milhões de oócitos. No momento da menarca, seu número diminui para 300 mil. Durante todo o período de vida reprodutiva, atinge a maturidade e ovula não mais de 500 folículos. O crescimento inicial dos folículos não depende da estimulação da FSH, é limitado e a atresia ocorre rapidamente. Acredita-se que, em lugar de hormônios esteróides, os péptidos autocrinos / paracrinos locais são o principal regulador do crescimento e da atresia dos folículos primários. Acredita-se que o processo de crescimento e atresia dos folículos não seja interrompido por nenhum processo fisiológico. Este processo continua em todas as idades, incluindo o período intra-uterino e a menopausa, é interrompido pela gravidez, ovulação e anovulação. O mecanismo que desencadeia o crescimento dos folículos e seu número em cada ciclo específico ainda não está claro.
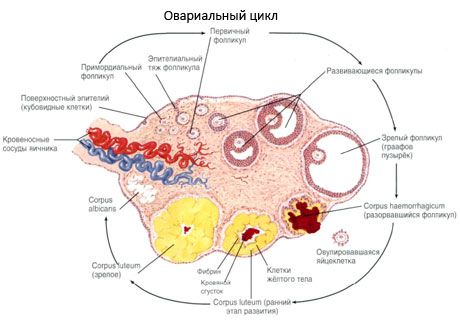
No seu desenvolvimento, o folículo passa por vários estágios de desenvolvimento. As células germinativas primordiais são originárias do endoderma do saco vitelino, alantoides e migram para a área genital do embrião na quinta e sexta semana de gravidez. Como resultado da rápida divisão mitótica, que dura de 6-8 semanas a 16-20 semanas de gravidez, são formados até 6-7 milhões de oócitos nos ovários do embrião, rodeados por uma fina camada de células da granulosa.
O folículo pré-antral - o oócito é cercado por uma membrana (Zona Pellucida). As células de granulosa que cercam o oócito começam a proliferar, seu crescimento depende das gonadotropinas e correlaciona-se com o nível de estrogênios. As células da granulosa são alvo da FSH. No estágio do folículo pré-antral, as células da granulosa são capazes de sintetizar três classes de esteróides: induz preferencialmente a atividade da aromatase, principal enzima que converte andrógenos em estradiol. Acredita-se que o estradiol é capaz de aumentar o número de seus próprios receptores, proporcionando um efeito mitogênico direto nas células da granulosa, independentemente da FSH. É considerado um fator paracrino que melhora os efeitos da FSH, incluindo a ativação dos processos de aromatização.
Os receptores da FSH aparecem nas membranas das células da granulosa assim que o crescimento do folículo começa. A redução ou aumento da FSH leva a uma mudança no número de receptores. Essa ação da FSH é modulada por fatores de crescimento. A FSH atua através da proteína G, do sistema adenilato-ciclase, embora a esteroidogênese no folículo seja principalmente regulada pela FSH, muitos fatores estão envolvidos neste processo: canais de iões, receptores de tirosina quinase, sistema de fosfolipase de mensageiros secundários.
O papel dos andrógenos no desenvolvimento inicial do folículo é complexo. As células de granulosa possuem receptores de andrógenos. Eles não são apenas um substrato para a aromatização induzida por FSH nos estrogénios, mas podem melhorar o processo de aromatização em baixas concentrações. Quando o nível de androgénios aumenta as células da granulosa pré-antrais preferencialmente seleccionado caminho sem aromatização em estrógenos e maneira mais simples para a conversão através de androgénios 5a-redutase em um androgénio em desenvolvimento, que não pode ser convertida em estrogénio, e a actividade da aromatase, assim inibida. Este processo também inibe FSH e a formação de receptores LH, interrompendo assim o desenvolvimento do folículo.
O processo de aromatização, um folículo com alto nível de andrógenos, sofre processos de atresia. O crescimento eo desenvolvimento do folículo dependem da sua capacidade de converter andrógenos em estrogênios.
Na presença de FSH, a substância dominante do líquido folicular é estrogênios. Na ausência de FSH - andrógenos. LH é normal no fluido folicular até o meio do ciclo. Logo que o aumento do nível de actividade mitótica das células da granulosa de LH diminui, e as mudanças degenerativas apareça aumento do nível de androgénio nos níveis folículo esteróides no fluido folicular do que no plasma e reflecte a actividade dos ovários funcional: As células da granulosa e tecais. Se o único alvo para a FSH são células da granulosa, LH tem muitos alvos - estas são células, células estromáticas e lúteas e células da granulosa. A capacidade de esteroidogênese possui células de granulosa e teka, mas a atividade da aromatase predomina nas células da granulosa.
Em resposta à LH, as células teka produzem andrógenos, que então, através da aromatização induzida por FSH, são transformados por células de granulosa em estrogênios.
À medida que o folículo de células da teca começam a expressar os genes para o receptor de LH s P450 e desidrogenase 3beta-hidroxiesteróide, factor de crescimento semelhante à insulina (IGF-1) sinergicamente com LH para aumentar a expressão do gene, mas não estimulam a esteroidogénese.
A esteroidogênese do ovário é sempre dependente de LH. À medida que o folículo cresce, as células atuais expressam a enzima P450c17, que forma andrógeno a partir do colesterol. As células da granulosa não possuem essa enzima e são dependentes das células atuais na produção de estrogênios dos andrógenos. Ao contrário da esteroidogênese - a foliculogênese depende da FSH. À medida que o folículo e aumentar o nível de estrogénio entra em mecanismo de feedback acção - inibiu a produção de FSH, que por sua vez leva a uma diminuição no folículo actividade da aromatase e, em última análise, para a atresia folicular através de apoptose (morte celular programada).
O mecanismo de feedback dos estrogênios e FSH inibe o desenvolvimento de folículos que começaram a crescer, mas não o folículo dominante. O folículo dominante contém mais receptores de FSH que suportam a proliferação de células da granulosa e a aromatização dos andrógenos nos estrogênios. Além disso, a via paracrina e autocrina atua como um coordenador importante para o desenvolvimento do folículo antral.
Uma parte integrante do regulador autocrino / paracrino são péptidos (inibina, activina, folicatina), que são sintetizados por células da granulosa em resposta à ação da FSH e entram no líquido folicular. A inibina reduz a secreção de FSH; A activina estimula a liberação de FSH da glândula pituitária e melhora a ação da FSH no ovário; A folistatina suprime a atividade da FSH, possivelmente devido à ligação da activina. Após a ovulação e desenvolvimento do corpo amarelo, a inibina está sob o controle de LH.
O crescimento e a diferenciação das células do ovário são influenciados por fatores de crescimento semelhantes a insulina (IGE). O IGF-1 atua sobre as células da granulosa, causando um aumento do monofosfato de adenosina cíclica (cAMP), progesterona, oxitocina, proteoglicano e inibina.
IGF-1 atua sobre células teka, causando um aumento na produção de andrógenos. As células Teka, por sua vez, produzem o fator de necrose tumoral (TNF) e o fator de crescimento epidérmico (EGF), que também são regulados pela FSH.
EGF estimula a proliferação de células da granulosa. O IGF-2 é o principal fator de crescimento do fluido folicular, também detectou IGF-1, TNF-a, TNF-3 e EGF.
A violação da regulação paracrina e / ou autócrina da função ovariana parece desempenhar um papel nos distúrbios dos processos de ovulação e na formação de ovários policísticos.
À medida que o folículo antral cresce, o teor de estrogênios no fluido folicular aumenta. No auge de seu aumento nas células da granulosa, aparecem receptores para LH, a luteinização das células da granulosa ocorre e a produção de progesterona aumenta. Assim, durante o período pré-ovulatório, um aumento na produção de estrogênios provoca o aparecimento de receptores de LH, LH, por sua vez, provoca a luteinização de células da granulosa e produção de progesterona. O aumento da progesterona reduz o nível de estrogênios, o que, aparentemente, causa o segundo pico de FSH no meio do ciclo.
Acredita-se que a ovulação ocorre 10-12 horas após o pico de LH e 24-36 horas após o pico de estradiol. Acredita-se que a LH estimula a redução do oócito, a luteinização de células da granulosa, a síntese de progesterona e prostaglandina no folículo.
A progesterona aumenta a atividade das enzimas proteolíticas, juntamente com a prostaglandina envolvida na ruptura da parede do folículo. Pico de FSH induzido por progesterona, permite ao oócito sair do folículo convertendo o plasminogênio em uma enzima proteolítica-plasmina, fornece um número suficiente de receptores de LH para o desenvolvimento normal da fase lútea.
Dentro de 3 dias após a ovulação, as células da granulosa aumentam, as vacuolas características preenchidas com pigmento, a luteína, aparecem nelas. As células Teka-lúteas diferenciam-se do teki e do estroma e tornam-se parte do corpo amarelo. Muito rapidamente, sob a influência de fatores de angiogênese, o desenvolvimento de capilares que penetram no corpo amarelo está progredindo e, com a melhora da vascularização, aumenta a produção de progesterona e estrogênios. A atividade da esteroidogênese e a duração do corpo amarelo são determinadas pelo nível de LH. O corpo amarelo não é uma entidade celular homogênea. A adição de células 2 tipos lútea que contém células endoteliais, macrófagos, fibroblastos e outros. Lútea células grandes produzir péptidos (relaxina, oxitocina) e são mais activos na esteroidogénese actividade mais aromatase e um grande síntese de progesterona do que as pequenas células.
O pico da progesterona é observado no 8º dia após o pico da LG. Observou-se que a progesterona e estradiol na fase lútea são segregados esporadicamente em correlação com a saída de pulso de LH. Com a formação de um corpo amarelo, o controle sobre a produção de inibina passa de FSH para LH. Ingibin aumenta com o aumento do estradiol até o pico de LH e continua a aumentar após o pico de LH, embora o nível de estrogênios diminua. Embora a inibina e o estradiol sejam segregados por células da granulosa, eles são regulados de diferentes maneiras. A diminuição da inibina no final da fase lútea contribui para um aumento da FSH para o próximo ciclo.
O corpo amarelo muito rapidamente - no dia 9-11 após a ovulação diminuir.
O mecanismo de degeneração não é claro e não está relacionado com o papel líuteolático dos estrogênios ou com o mecanismo vinculado ao receptor, como observado no endométrio. Há outra explicação para o papel dos estrogénios produzidos pelo corpo amarelo. Sabe-se que, para a síntese de receptores de progesterona no endométrio, são necessários estrogênios. Os estrogénios da fase lútea são provavelmente necessários para alterações relacionadas à progesterona no endométrio após a ovulação. O desenvolvimento inadequado dos receptores de progesterona, como resultado do teor inadequado de estrogênio, é provavelmente um mecanismo adicional de infertilidade e perda precoce da gravidez, outra forma de inferioridade da fase lútea. Acredita-se que a vida útil do corpo amarelo esteja ajustada no momento da ovulação. E certamente será regredido se a gonadotropina coriônica não for apoiada em conexão com a gravidez. Assim, a regressão do corpo amarelo leva a uma diminuição dos níveis de estradiol, progesterona e inibina. A inibina de redução remove o seu efeito inibitório na FSH; A redução do estradiol e da progesterona permite recuperar muito rapidamente a secreção de GnRH e remover o mecanismo de feedback da hipófise. A redução da inibina e estradiol, juntamente com o aumento da GnRH, leva à prevalência de FSH sobre LH. Um aumento na FSH leva ao crescimento dos folículos com a escolha subseqüente de um folículo dominante e um novo ciclo começa, no caso de a gravidez não ocorrer. Os hormônios esteróides desempenham um papel de liderança na biologia reprodutiva e na fisiologia geral. Eles determinam o fenótipo de uma pessoa, afetam o sistema cardiovascular, o metabolismo dos ossos, a pele, o bem-estar geral do corpo e desempenham um papel fundamental na gravidez. A ação dos hormônios esteróides reflete os mecanismos intracelulares e genéticos que são necessários para transferir o sinal extracelular para o núcleo da célula para induzir uma resposta fisiológica.
Os estrogénios difundem através da membrana celular e se ligam aos receptores localizados no núcleo da célula. O complexo receptor-esteróide então se liga ao DNA. Nas células alvo, essas interações levam à expressão de genes, a síntese de proteínas, a uma função específica de células e tecidos.