Médico especialista do artigo

Novas publicações
glândula pituitária
Última revisão: 04.07.2025

Todo o conteúdo do iLive é medicamente revisado ou verificado pelos fatos para garantir o máximo de precisão factual possível.
Temos diretrizes rigorosas de fornecimento e vinculamos apenas sites de mídia respeitáveis, instituições de pesquisa acadêmica e, sempre que possível, estudos médicos revisados por pares. Observe que os números entre parênteses ([1], [2], etc.) são links clicáveis para esses estudos.
Se você achar que algum dos nossos conteúdos é impreciso, desatualizado ou questionável, selecione-o e pressione Ctrl + Enter.
A hipófise (glândula pituitária) está localizada na fossa hipofisária da sela túrcica do osso esfenoide e é separada da cavidade craniana por um processo da dura-máter do cérebro, formando o diafragma da sela. Através da abertura neste diafragma, a hipófise se conecta ao infundíbulo do hipotálamo do diencéfalo. O tamanho transversal da hipófise é de 10 a 17 mm, o anteroposterior é de 5 a 15 mm e o vertical é de 5 a 10 mm. A massa da hipófise nos homens é de aproximadamente 0,5 g, nas mulheres - 0,6 g. A hipófise é revestida externamente por uma cápsula.
De acordo com o desenvolvimento da hipófise a partir de dois rudimentos diferentes, dois lobos são distinguidos no órgão - o anterior e o posterior. A adeno-hipófise, ou lobo anterior (adeno-hipófise, s.lobus anterior), é maior, constituindo 70-80% da massa total da hipófise. É mais densa que o lobo posterior. No lobo anterior, distingue-se uma parte distal (pars distalis), que ocupa a parte anterior da fossa hipofisária, uma parte intermediária (pars intermedia), localizada na fronteira com o lobo posterior, e uma parte tuberosa (pars tuberalis), que sobe e se conecta com o infundíbulo do hipotálamo. Devido à abundância de vasos sanguíneos, o lobo anterior tem uma cor amarelo-pálida com um tom avermelhado. O parênquima da hipófise anterior é representado por vários tipos de células glandulares, entre os filamentos dos quais estão localizados capilares sanguíneos sinusoidais. Metade (50%) das células da adeno-hipófise são adenócitos cromofílicos, que apresentam grânulos finos em seu citoplasma que se coram bem com sais de crômio. São eles: adenócitos acidófilos (40% de todas as células da adeno-hipófise) e adenócitos basófilos (10%). Os adenócitos basófilos incluem endocrinócitos gonadotrópicos, corticotrópicos e tireotrópicos. Os adenócitos cromofóbicos são pequenos, possuem um núcleo grande e uma pequena quantidade de citoplasma. Essas células são consideradas precursoras dos adenócitos cromofílicos. Os outros 50% das células da adeno-hipófise são adenócitos cromofóbicos.
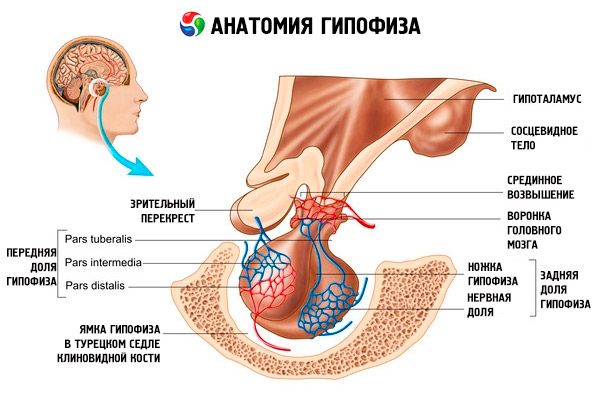
A neuro-hipófise, ou lobo posterior (neuro-hipófise, s.lobus posterior), consiste no lobo neural (lobus nervosus), localizado na parte posterior da fossa hipofisária, e no funil (infundíbulo), localizado atrás da parte tuberosa da adeno-hipófise. O lobo posterior da hipófise é formado por células neurogliais (células hipofisárias), fibras nervosas que vão dos núcleos neurossecretores do hipotálamo à neuro-hipófise e corpúsculos neurossecretores.
A hipófise, por meio de fibras nervosas (vias) e vasos sanguíneos, está funcionalmente conectada ao hipotálamo do diencéfalo, que regula a atividade da hipófise. A hipófise e o hipotálamo, juntamente com suas conexões neuroendócrinas, vasculares e nervosas, são geralmente considerados o sistema hipotálamo-hipofisário.
Os hormônios das glândulas pituitárias anterior e posterior afetam muitas funções do corpo, principalmente por meio de outras glândulas endócrinas. Na glândula pituitária anterior, os adenócitos acidófilos (células alfa) produzem o hormônio somatotrópico (hormônio do crescimento), que participa da regulação dos processos de crescimento e desenvolvimento em organismos jovens. Os endocrinócitos corticotrópicos secretam o hormônio adrenocorticotrófico (ACTH), que estimula a secreção de hormônios esteroides pelas glândulas suprarrenais. Os endocrinócitos tireotrópicos secretam o hormônio tireotrópico (TSH), que afeta o desenvolvimento da glândula tireoide e ativa a produção de seus hormônios. Os hormônios gonadotrópicos: folículo-estimulante (FSH), luteinizante (LH) e prolactina - afetam a maturação sexual do corpo, regulam e estimulam o desenvolvimento dos folículos ovarianos, a ovulação, o crescimento das glândulas mamárias e a produção de leite nas mulheres, e o processo de espermatogênese nos homens. Esses hormônios são produzidos por adenócitos basofílicos (células beta). Fatores lipotrópicos da hipófise também são secretados por ela, afetando a mobilização e a utilização de gorduras no corpo. Na parte intermediária do lobo anterior, é produzido o hormônio estimulador dos melanócitos, que controla a formação de pigmentos – melaninas – no corpo.
As células neurossecretoras dos núcleos supraóptico e paraventricular no hipotálamo produzem vasopressina e ocitocina. Esses hormônios são transportados para as células da hipófise posterior ao longo dos axônios que compõem o trato hipotálamo-hipofisário. A partir da hipófise posterior, essas substâncias entram na corrente sanguínea. O hormônio vasopressina tem efeito vasoconstritor e antidiurético, sendo também chamado de hormônio antidiurético (ADH). A ocitocina tem efeito estimulante sobre a contratilidade dos músculos uterinos, aumenta a secreção de leite pela glândula mamária lactante, inibe o desenvolvimento e a função do corpo lúteo e afeta as alterações no tônus da musculatura lisa (não estriada) do trato gastrointestinal.
Desenvolvimento da glândula pituitária
O lobo anterior da hipófise desenvolve-se a partir do epitélio da parede dorsal da cavidade oral, na forma de uma protuberância em forma de anel (bolsa de Rathke). Essa protrusão ectodérmica cresce em direção à base do futuro terceiro ventrículo. Em direção a ela, a partir da superfície inferior da segunda vesícula cerebral (futura base do terceiro ventrículo), cresce um processo, a partir do qual se desenvolvem o tubérculo cinzento do infundíbulo e o lobo posterior da hipófise.
Vasos e nervos da glândula pituitária
Das artérias carótidas internas e dos vasos do círculo arterial do cérebro, as artérias pituitárias superior e inferior são direcionadas para a hipófise. As artérias pituitárias superiores vão para o núcleo cinzento e o infundíbulo do hipotálamo, anastomosam-se entre si e formam capilares que penetram no tecido cerebral - a rede hemocapilar primária. A partir das alças longas e curtas dessa rede, formam-se as veias porta, que são direcionadas para o lobo anterior da hipófise. No parênquima do lobo anterior da hipófise, essas veias se dividem em capilares sinusoidais largos, formando a rede hemocapilar secundária. O lobo posterior da hipófise é suprido com sangue principalmente pela artéria pituitária inferior. Existem longas anastomoses arteriais entre as artérias pituitárias superior e inferior. A saída do sangue venoso da rede hemocapilar secundária é realizada através de um sistema de veias que desembocam nos seios cavernosos e intercavernosos da dura-máter do cérebro.
As fibras simpáticas que entram no órgão juntamente com as artérias participam da inervação da hipófise. As fibras nervosas simpáticas pós-ganglionares partem do plexo da artéria carótida interna. Além disso, numerosas terminações dos prolongamentos das células neurossecretoras localizadas nos núcleos do hipotálamo são encontradas no lobo posterior da hipófise.
Características da glândula pituitária relacionadas à idade
O peso médio da hipófise em recém-nascidos atinge 0,12 g. O peso do órgão dobra aos 10 anos de idade e triplica aos 15. Aos 20 anos, o peso da hipófise atinge seu máximo (530-560 mg) e permanece praticamente inalterado nos períodos subsequentes. Após os 60 anos, observa-se uma ligeira diminuição no peso dessa glândula endócrina.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hormônios da hipófise
A unidade da regulação nervosa e hormonal no corpo é garantida pela estreita conexão anatômica e funcional entre a hipófise e o hipotálamo. Este complexo determina o estado e o funcionamento de todo o sistema endócrino.
A principal glândula endócrina que produz uma série de hormônios peptídicos que regulam diretamente a função das glândulas periféricas é a hipófise. É uma formação cinza-avermelhada em forma de feijão coberta por uma cápsula fibrosa pesando 0,5-0,6 g. Varia ligeiramente dependendo do sexo e da idade de uma pessoa. A divisão geralmente aceita da hipófise em dois lobos que diferem em desenvolvimento, estrutura e função permanece: o distal anterior - adeno-hipófise e o posterior - neuro-hipófise. O primeiro compõe cerca de 70% da massa total da glândula e é convencionalmente dividido nas partes distal, infundibular e intermediária, o segundo - na parte posterior, ou lobo, e o pedúnculo hipofisário. A glândula está localizada na fossa hipofisária da sela túrcica do osso esfenoide e está conectada ao cérebro através do pedúnculo. A parte superior do lobo anterior é coberta pelo quiasma óptico e tratos ópticos. O suprimento sanguíneo para a hipófise é muito abundante e é realizado por ramos da artéria carótida interna (as artérias hipofisárias superior e inferior), bem como por ramos do círculo arterial do cérebro. As artérias hipofisárias superiores participam do suprimento sanguíneo da adeno-hipófise, e as inferiores, da neuro-hipófise, contatando as terminações neurossecretoras dos axônios dos núcleos de grandes células do hipotálamo. As primeiras entram na eminência mediana do hipotálamo, onde se dispersam em uma rede capilar (o plexo capilar primário). Esses capilares (com os quais os terminais dos axônios das pequenas células neurossecretoras do hipotálamo mediobasal entram em contato) se acumulam nas veias porta que descem ao longo do pedúnculo hipofisário até o parênquima da adeno-hipófise, onde novamente se dividem em uma rede de capilares sinusoidais (o plexo capilar secundário). Assim, o sangue, tendo passado previamente pela eminência mediana do hipotálamo, onde é enriquecido com hormônios adeno-hipofisótropos hipotalâmicos (hormônios liberadores), entra na adeno-hipófise.
O fluxo de sangue saturado com hormônios adeno-hipofisários de numerosos capilares do plexo secundário é realizado através do sistema venoso, que por sua vez desemboca nos seios venosos da dura-máter e, em seguida, na corrente sanguínea geral. Assim, o sistema porta da hipófise, com fluxo sanguíneo descendente do hipotálamo, é um componente morfofuncional do complexo mecanismo de controle neuro-humoral das funções tróficas da adeno-hipófise.
A hipófise é inervada por fibras simpáticas que acompanham as artérias hipofisárias. Elas se originam de fibras pós-ganglionares que passam pelo plexo carotídeo interno, conectadas aos gânglios cervicais superiores. Não há inervação direta da adeno-hipófise pelo hipotálamo. O lobo posterior recebe fibras nervosas dos núcleos neurossecretores do hipotálamo.
A adeno-hipófise é uma formação muito complexa em sua arquitetura histológica. Ela possui dois tipos de células glandulares: cromofóbicas e cromofílicas. Estas últimas, por sua vez, são divididas em acidófilas e basofílicas (uma descrição histológica detalhada da hipófise é fornecida na seção correspondente do manual). No entanto, deve-se notar que os hormônios produzidos pelas células glandulares que compõem o parênquima da adeno-hipófise, devido à diversidade destas últimas, são, em certa medida, diferentes em sua natureza química, e a estrutura fina das células secretoras deve corresponder às características de biossíntese de cada uma delas. Às vezes, porém, formas transitórias de células glandulares capazes de produzir vários hormônios podem ser observadas na adeno-hipófise. Há evidências de que o tipo de células glandulares da adeno-hipófise nem sempre é determinado geneticamente.
Abaixo do diafragma da sela túrcica encontra-se a parte em forma de funil do lobo anterior. Ela envolve o pedúnculo da hipófise, em contato com o tubérculo cinzento. Essa parte da adeno-hipófise é caracterizada pela presença de células epiteliais e abundante suprimento sanguíneo. Também é hormonalmente ativa.
A parte intermediária (meio) da glândula pituitária consiste em várias camadas de grandes células basofílicas secretoras ativas.
A hipófise desempenha diversas funções por meio de seus hormônios. Seu lobo anterior produz hormônios adrenocorticotróficos (ACTH), estimulantes da tireoide (TSH), folículo-estimulantes (FSH), luteinizantes (LH), lipotrópicos, além do hormônio do crescimento somatotrópico (STO) e da prolactina. No lobo intermediário, o hormônio melanócito-estimulante (MSH) é sintetizado, e no lobo posterior, a vasopressina e a ocitocina se acumulam.
ACTH
Os hormônios hipofisários são um grupo de hormônios proteicos, peptídicos e glicoproteínas. Dos hormônios da hipófise anterior, o ACTH é o mais estudado. É produzido por células basofílicas. Sua principal função fisiológica é estimular a biossíntese e a secreção de hormônios esteroides pelo córtex adrenal. O ACTH também exibe atividade lipotrópica e estimulante de melanócitos. Em 1953, foi isolado em sua forma pura. Posteriormente, sua estrutura química foi estabelecida, consistindo de 39 resíduos de aminoácidos em humanos e em vários mamíferos. O ACTH não possui especificidade de espécie. Atualmente, a síntese química do próprio hormônio e de vários fragmentos de sua molécula, mais ativos do que os hormônios naturais, foi realizada. A estrutura do hormônio possui duas seções da cadeia peptídica, uma das quais garante a detecção e a ligação do ACTH ao receptor, e a outra proporciona um efeito biológico. Aparentemente, ele se liga ao receptor de ACTH por meio da interação das cargas elétricas do hormônio e do receptor. O papel do efetor biológico do ACTH é desempenhado pelo fragmento da molécula 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
A atividade estimulante dos melanócitos do ACTH deve-se à presença na molécula da região N-terminal, composta por 13 resíduos de aminoácidos e que repete a estrutura do hormônio alfa-melanócito estimulante. Essa região também contém um heptapeptídeo presente em outros hormônios hipofisários e que possui algumas atividades adrenocorticotrópicas, estimulantes dos melanócitos e lipotrópicas.
O momento-chave na ação do ACTH é considerado a ativação da enzima proteína quinase no citoplasma, com a participação do AMPc. A proteína quinase fosforilada ativa a enzima esterase, que converte ésteres de colesterol em uma substância livre em gotículas de gordura. A proteína sintetizada no citoplasma como resultado da fosforilação dos ribossomos estimula a ligação do colesterol livre ao citocromo P-450 e sua transferência das gotículas lipídicas para a mitocôndria, onde estão presentes todas as enzimas que garantem a conversão do colesterol em corticosteroides.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Hormônio estimulante da tireoide
TSH - tireotropina - o principal regulador do desenvolvimento e funcionamento da glândula tireoide, dos processos de síntese e secreção dos hormônios tireoidianos. Essa proteína complexa - glicoproteína - consiste em subunidades alfa e beta. A estrutura da primeira subunidade coincide com a subunidade alfa do hormônio luteinizante. Além disso, ela coincide amplamente em diferentes espécies animais. A sequência de resíduos de aminoácidos na subunidade beta do TSH humano foi decifrada e consiste em 119 resíduos de aminoácidos. Pode-se notar que as subunidades beta do TSH humano e bovino são semelhantes em muitos aspectos. As propriedades biológicas e a natureza da atividade biológica dos hormônios glicoproteicos são determinadas pela subunidade beta. Ela também garante a interação do hormônio com receptores em vários órgãos-alvo. No entanto, a subunidade beta na maioria dos animais exibe atividade específica somente após se combinar com a subunidade alfa, que atua como uma espécie de ativador do hormônio. Este último induz, com igual probabilidade, atividades luteinizante, folículo-estimulante e tireotrópica, determinadas pelas propriedades da subunidade beta. A similaridade encontrada permite concluir que esses hormônios surgiram no processo de evolução a partir de um precursor comum: a subunidade beta, que também determina as propriedades imunológicas dos hormônios. Supõe-se que a subunidade alfa proteja a subunidade beta da ação de enzimas proteolíticas e também facilite seu transporte da hipófise para os órgãos-alvo periféricos.
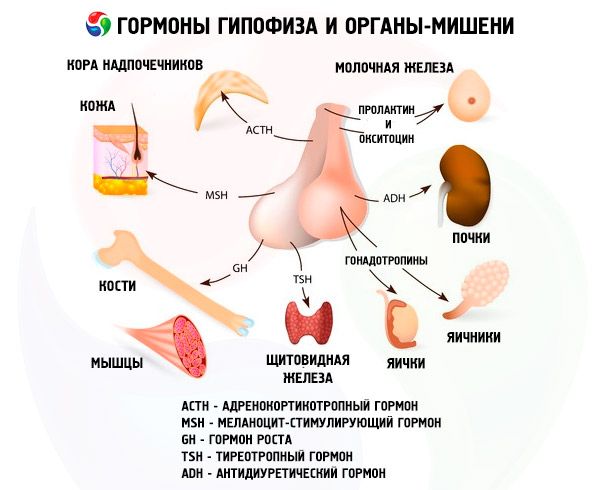
Hormônios gonadotrópicos
As gonadotrofinas estão presentes no corpo na forma de LH e FSH. A função desses hormônios geralmente se resume a garantir os processos reprodutivos em indivíduos de ambos os sexos. Assim como o TSH, elas são proteínas complexas – glicoproteínas. O FSH induz a maturação dos folículos nos ovários das mulheres e estimula a espermatogênese nos homens. O LH causa a ruptura folicular nas mulheres com a formação do corpo lúteo e estimula a secreção de estrogênios e progesterona. Nos homens, o mesmo hormônio acelera o desenvolvimento do tecido intersticial e a secreção de andrógenos. Os efeitos das gonadotrofinas são interdependentes e ocorrem de forma sincronizada.
A dinâmica da secreção de gonadotrofinas nas mulheres muda durante o ciclo menstrual e tem sido estudada em detalhes suficientes. Na fase pré-ovulatória (folicular) do ciclo, o conteúdo de LH está em um nível bastante baixo, enquanto o de FSH está aumentado. À medida que o folículo amadurece, a secreção de estradiol aumenta, o que promove o aumento da produção de gonadotrofinas pela hipófise e a ocorrência de ciclos de LH e FSH, ou seja, os esteroides sexuais estimulam a secreção de gonadotrofinas.
Atualmente, a estrutura do LH foi determinada. Assim como o TSH, ele consiste em duas subunidades: a e b. A estrutura da subunidade alfa do LH em diferentes espécies animais coincide amplamente, correspondendo à estrutura da subunidade alfa do TSH.
A estrutura da subunidade beta do LH difere significativamente da estrutura da subunidade beta do TSH, embora possua quatro seções idênticas da cadeia peptídica, compostas por 4 a 5 resíduos de aminoácidos. No TSH, elas estão localizadas nas posições 27 a 31, 51 a 54, 65 a 68 e 78 a 83. Como a subunidade beta do LH e do TSH determina a atividade biológica específica dos hormônios, pode-se presumir que seções homólogas na estrutura do LH e do TSH devem garantir a conexão das subunidades beta com a subunidade alfa, e seções com estruturas diferentes devem ser responsáveis pela especificidade da atividade biológica dos hormônios.
O LH nativo é muito estável à ação de enzimas proteolíticas, mas a subunidade beta é rapidamente clivada pela quimotripsina, e a subunidade a é difícil de ser hidrolisada pela enzima, ou seja, ela desempenha um papel protetor, impedindo que a quimotripsina acesse as ligações peptídicas.
Quanto à estrutura química do FSH, os pesquisadores ainda não obtiveram resultados definitivos. Assim como o LH, o FSH consiste em duas subunidades, mas a subunidade beta do FSH difere da subunidade beta do LH.
Prolactina
Outro hormônio, a prolactina (hormônio lactogênico), desempenha um papel ativo nos processos reprodutivos. As principais propriedades fisiológicas da prolactina em mamíferos se manifestam na forma de estimulação do desenvolvimento das glândulas mamárias e da lactação, no crescimento das glândulas sebáceas e dos órgãos internos. Ela promove a manifestação do efeito dos esteroides nas características sexuais secundárias em machos, estimula a atividade secretora do corpo lúteo em camundongos e ratos e participa da regulação do metabolismo da gordura. Muita atenção tem sido dada à prolactina nos últimos anos como reguladora do comportamento materno; tal polifuncionalidade é explicada por seu desenvolvimento evolutivo. É um dos antigos hormônios hipofisários e é encontrado até mesmo em anfíbios. Atualmente, a estrutura da prolactina em algumas espécies de mamíferos foi completamente decifrada. No entanto, até recentemente, os cientistas expressaram dúvidas sobre a existência de tal hormônio em humanos. Muitos acreditavam que sua função era desempenhada pelo hormônio do crescimento. Agora, evidências convincentes foram obtidas da presença de prolactina em humanos e sua estrutura foi parcialmente decifrada. Os receptores de prolactina se ligam ativamente ao hormônio do crescimento e ao lactogênio placentário, indicando um único mecanismo de ação dos três hormônios.
Somatotropina
O hormônio do crescimento, somatotropina, tem um espectro de ação ainda mais amplo do que a prolactina. Assim como a prolactina, é produzido pelas células acidófilas da adeno-hipófise. O STH estimula o crescimento esquelético, ativa a biossíntese de proteínas, tem efeito mobilizador de gordura e promove o aumento do tamanho corporal. Além disso, coordena os processos metabólicos.
A participação do hormônio neste último é confirmada pelo fato de um aumento acentuado de sua secreção pela hipófise, por exemplo, quando o nível de açúcar no sangue diminui.
A estrutura química deste hormônio humano está agora totalmente estabelecida – 191 resíduos de aminoácidos. Sua estrutura primária é semelhante à da somatomamotropina coriônica ou do lactogênio placentário. Esses dados indicam uma proximidade evolutiva significativa entre os dois hormônios, embora apresentem diferenças na atividade biológica.
É necessário enfatizar a alta especificidade do hormônio em questão para cada espécie – por exemplo, o STH de origem animal é inativo em humanos. Isso se explica tanto pela reação entre os receptores de STH de humanos e animais quanto pela própria estrutura do hormônio. Atualmente, estudos estão em andamento para identificar centros ativos na complexa estrutura do STH que exibem atividade biológica. Fragmentos individuais da molécula que exibem outras propriedades estão sendo estudados. Por exemplo, após a hidrólise do STH humano pela pepsina, um peptídeo composto por 14 resíduos de aminoácidos e correspondente à seção 31-44 da molécula foi isolado. Não apresentou efeito de crescimento, mas excedeu significativamente o hormônio nativo em atividade lipotrópica. O hormônio do crescimento humano, diferentemente do hormônio similar de animais, possui atividade lactogênica significativa.
A adeno-hipófise sintetiza muitas substâncias peptídicas e proteicas com efeito mobilizador de gordura, e os hormônios trópicos hipofisários – ACTH, STH, TSH e outros – têm efeito lipotrópico. Nos últimos anos, os hormônios beta e y-lipotrópicos (LPG) têm sido particularmente enfatizados. As propriedades biológicas do beta-LPG têm sido estudadas detalhadamente; além da atividade lipotrópica, ele também possui efeito estimulante de melanócitos, estimulante de corticotropina e hipocalcêmico, além de produzir um efeito semelhante ao da insulina.
Atualmente, a estrutura primária do LPG ovino (90 resíduos de aminoácidos) e dos hormônios lipotrópicos de suínos e bovinos foi decifrada. Esse hormônio possui especificidade específica para cada espécie, embora a estrutura da região central do beta-LPG seja a mesma em diferentes espécies. Ela determina as propriedades biológicas do hormônio. Um dos fragmentos dessa região é encontrado na estrutura do alfa-MSH, beta-MSH, ACTH e beta-LPG. Sugere-se que esses hormônios tenham surgido do mesmo precursor no processo evolutivo. O γ-LPG possui uma atividade lipotrópica mais fraca que o beta-LPG.
Hormônio estimulante de melanócitos
Este hormônio, sintetizado no lobo intermediário da hipófise, estimula a biossíntese do pigmento da pele melanina. Em sua função biológica, promove um aumento no tamanho e no número de células pigmentares (melanócitos) na pele de anfíbios. Essas qualidades do MSH são utilizadas em testes biológicos do hormônio. Existem dois tipos do hormônio: alfa-MSH e beta-MSH. Foi demonstrado que o alfa-MSH não possui especificidade de espécie e possui a mesma estrutura química em todos os mamíferos. Sua molécula é uma cadeia peptídica composta por 13 resíduos de aminoácidos. O beta-MSH, por outro lado, possui especificidade de espécie e sua estrutura difere em diferentes animais. Na maioria dos mamíferos, a molécula de beta-MSH consiste em 18 resíduos de aminoácidos e, somente em humanos, ela se estende da extremidade do aminoácido por quatro resíduos de aminoácidos. Deve-se notar que o alfa-MSH possui alguma atividade adrenocorticotrópica e seu efeito no comportamento de animais e humanos já foi comprovado.
Ocitocina e vasopressina
A vasopressina e a ocitocina, sintetizadas no hipotálamo, acumulam-se no lobo posterior da hipófise: a vasopressina nos neurônios do núcleo supraóptico e a ocitocina no núcleo paraventricular. Em seguida, são transferidas para a hipófise. Deve-se enfatizar que o precursor do hormônio vasopressina é sintetizado primeiro no hipotálamo. Ao mesmo tempo, as proteínas neurofisina tipos 1 e 2 são produzidas ali. A primeira se liga à ocitocina e a segunda à vasopressina. Esses complexos migram na forma de grânulos neurossecretores no citoplasma ao longo do axônio e alcançam o lobo posterior da hipófise, onde as fibras nervosas terminam na parede vascular e o conteúdo dos grânulos entra no sangue. A vasopressina e a ocitocina são os primeiros hormônios hipofisários com uma sequência de aminoácidos completamente estabelecida. Em sua estrutura química, são nonapeptídeos com uma ponte dissulfeto.
Os hormônios em questão produzem vários efeitos biológicos: estimulam o transporte de água e sais através das membranas, têm efeito vasopressor, aumentam as contrações da musculatura lisa do útero durante o parto e aumentam a secreção das glândulas mamárias. Deve-se notar que a vasopressina tem uma atividade antidiurética maior do que a ocitocina, enquanto esta última tem um efeito mais forte no útero e na glândula mamária. O principal regulador da secreção de vasopressina é o consumo de água; nos túbulos renais, ela se liga a receptores nas membranas citoplasmáticas com subsequente ativação da enzima adenilato ciclase neles. Diferentes partes da molécula são responsáveis pela ligação do hormônio ao receptor e pelo efeito biológico.
A hipófise, conectada através do hipotálamo a todo o sistema nervoso, une o sistema endócrino em um todo funcional, contribuindo para garantir a constância do ambiente interno do corpo (homeostase). Dentro do sistema endócrino, a regulação homeostática é realizada com base no princípio de retroalimentação entre o lobo anterior da hipófise e as glândulas "alvo" (tireoide, córtex adrenal, gônadas). O excesso do hormônio produzido pela glândula "alvo" inibe, e sua deficiência estimula, a secreção e a liberação do hormônio trópico correspondente. O hipotálamo está incluído no sistema de retroalimentação. É nele que se localizam as zonas receptoras sensíveis aos hormônios das glândulas "alvo". Ao se ligarem especificamente aos hormônios que circulam no sangue e alterarem a resposta dependendo da concentração hormonal, os receptores hipotalâmicos transmitem seu efeito aos centros hipotalâmicos correspondentes, que coordenam o trabalho da adeno-hipófise, liberando hormônios adeno-hipofisiotrópicos hipotalâmicos. Portanto, o hipotálamo deve ser considerado um cérebro neuroendócrino.