Médico especialista do artigo

Novas publicações
Modelos experimentais de osteoartrite
Última revisão: 07.07.2025

Todo o conteúdo do iLive é medicamente revisado ou verificado pelos fatos para garantir o máximo de precisão factual possível.
Temos diretrizes rigorosas de fornecimento e vinculamos apenas sites de mídia respeitáveis, instituições de pesquisa acadêmica e, sempre que possível, estudos médicos revisados por pares. Observe que os números entre parênteses ([1], [2], etc.) são links clicáveis para esses estudos.
Se você achar que algum dos nossos conteúdos é impreciso, desatualizado ou questionável, selecione-o e pressione Ctrl + Enter.
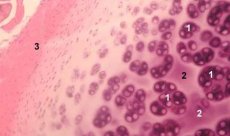
A cartilagem é um tecido altamente especializado que contém apenas um tipo de célula (condrócitos) e é caracterizada pela ausência de vasos sanguíneos e linfáticos. A cartilagem é nutrida principalmente pela absorção do líquido sinovial. O metabolismo dos condrócitos é regulado por uma série de fatores solúveis produzidos localmente pelos condrócitos e pelos tecidos circundantes. A função dos condrócitos também depende da composição do ambiente extracelular (tensão de oxigênio, concentração de íons, pH, etc.), da composição da MEC, da interação das células com a matriz e de sinais físicos. O principal objetivo da modelagem experimental é criar culturas no ambiente extracelular sem alterar o fenótipo das células maduras. O segundo objetivo é criar culturas para estudar a resposta prematura, tardia, de curto prazo ou prolongada dos condrócitos a sinais químicos e/ou físicos. Estudos in vitro também oferecem uma oportunidade para estudar o comportamento dos condrócitos na osteoartrose. O terceiro objetivo é desenvolver sistemas de cocultura que permitam estudar as interações de vários tecidos na articulação. A quarta tarefa é a preparação de implantes de cartilagem para transplante subsequente. E, finalmente, a quinta tarefa é o estudo de fatores de crescimento, citocinas ou agentes terapêuticos capazes de estimular a reparação e/ou inibir a reabsorção da cartilagem.
Nas últimas décadas, vários modelos de culturas de células de cartilagem articular foram criados, incluindo culturas em monocamada, culturas suspensas, culturas de condrócitos, explantes, coculturas e culturas de células imortais. Cada cultura tem suas próprias vantagens e desvantagens, e cada uma é adequada para estudar um aspecto específico do metabolismo de condrócitos. Assim, os explantes de cartilagem são um excelente modelo para estudar a renovação de elementos da matriz, o que requer receptores de superfície celular genuínos e interações célula-matriz e matriz-célula normais. Ao mesmo tempo, recomenda-se estudar depósitos de matriz ou mecanismos que regulam o metabolismo de condrócitos em uma cultura de células isoladas. Uma cultura em monocamada de baixa densidade é necessária para estudar o processo de diferenciação celular. Culturas suspensas em uma matriz natural ou sintética são um modelo para analisar a resposta adaptativa de condrócitos ao estresse mecânico.
Culturas de condrócitos
Vários pontos importantes devem ser levados em consideração ao selecionar tecido cartilaginoso para estudos in vitro. A composição da matriz e a atividade metabólica dos condrócitos variam entre as articulações, e esta última também depende da profundidade da localização dos condrócitos no tecido. Esses dados foram obtidos em vários experimentos nos quais subpopulações isoladas de condrócitos de zonas cartilaginosas de diferentes profundidades foram estudadas. Diversas diferenças morfológicas e bioquímicas foram encontradas entre condrócitos cultivados localizados nas camadas superficial e profunda da cartilagem articular. As células superficiais sintetizam uma matriz fibrilar esparsa e pobre em proteoglicanos, enquanto as células mais profundas produzem uma matriz rica em fibrilas e proteoglicanos. Além disso, as células superficiais produzem relativamente mais proteoglicanos pequenos não agregados e ácido hialurônico e relativamente menos agrecano e sulfato de queratana do que os condrócitos mais profundos. Outra importante característica distintiva do metabolismo de condrócitos isolados de zonas cartilaginosas de diferentes profundidades é a resposta a um estímulo exógeno. Segundo M. Aydelotte et al., os condrócitos bovinos da zona superficial da cartilagem foram mais sensíveis à IL-1 do que as células da zona profunda.
O comportamento celular também depende da localização do tecido. Condrócitos da cartilagem da costela e da orelha do mesmo animal respondem de forma diferente a fatores de crescimento, como o fator de crescimento de fibroblastos (FGF) e o TGF-beta. O FGF aumentou a incorporação de timidina, prolina e leucina em condrócitos de costela cultivados, mas não em condrócitos da orelha. O TGF-beta aumentou a incorporação de timidina em condrócitos da cartilagem da costela e da orelha, mas não teve efeito na incorporação de timidina e prolina em condrócitos da orelha. Células de cartilagem de áreas de alto estresse diferem daquelas de áreas de baixo estresse na cartilagem. Assim, condrócitos de cartilagem de joelho de carneiro maduro da região central da superfície articular da tíbia não coberta pelo menisco, que suporta a maior carga in vivo, sintetizam menos agrecano, mas mais decorina do que células das zonas cobertas pelo menisco. Os autores também enfatizam a importância do uso de cartilagem de zonas articulares idênticas ao estudar a função sintética das articulações.
O metabolismo dos condrócitos e sua resposta a fatores regulatórios também dependem significativamente da idade do doador, do seu desenvolvimento esquelético e da condição das articulações das quais as células são retiradas. Em condrócitos humanos, observa-se uma diminuição significativa na resposta proliferativa com a idade. A maior diminuição é observada em doadores com idade entre 40-50 anos e acima de 60 anos. Além disso, a gravidade da resposta proliferativa a fatores de crescimento (por exemplo, FGF e TGF-beta) diminui com o envelhecimento. Além das alterações quantitativas na proliferação de condrócitos, também há alterações qualitativas. Células de doadores jovens (10-20 anos) respondem melhor ao fator de crescimento derivado de plaquetas (PDGF) do que ao TGF-beta, enquanto o oposto é observado em células de doadores adultos. Vários mecanismos são usados para explicar as alterações dependentes da idade na função sintética dos condrócitos e sua resposta aos fatores de crescimento. Isso inclui uma diminuição no número e na afinidade dos receptores de células de superfície, alterações na síntese e bioatividade de fatores de crescimento e citocinas, e modificação de sinais pós-receptores.
A condição patológica das articulações também altera a morfologia e a atividade metabólica dos condrócitos. Assim, J. Kouri et al. (1996) identificaram três subpopulações de condrócitos na cartilagem na osteoartrose. Os condrócitos da parte superficial e superior do meio da cartilagem formam aglomerados e sintetizam uma quantidade maior de proteoglicanos e colágeno. O TGF-beta e o fator de crescimento semelhante à insulina (IGF) são capazes de estimular a síntese de proteoglicanos pelos condrócitos e neutralizar parcialmente os efeitos da IL-1 e do TNF-a. Explantes de cartilagem afetada pela osteoartrose e condrócitos isolados da cartilagem de um paciente com osteoartrose são mais sensíveis à estimulação do TGF-beta do que os condrócitos da cartilagem saudável. Essas diferenças estão provavelmente associadas a alterações fenotípicas nos condrócitos nas camadas superiores da cartilagem articular.
O isolamento de condrócitos individuais é obtido pelo tratamento sequencial da MEC com enzimas proteolíticas. Após sua liberação da MEC, as células isoladas são ideais para estudar a síntese de novo de componentes da matriz. Alguns autores usam apenas colagenase clostridial, enquanto outros pré-incubam a cartilagem com tripsina, pronase, DNase e/ou hialuronidase. O número de células isoladas depende das enzimas utilizadas. Assim, quando tratadas apenas com colagenase, 1,4-10 6 condrócitos podem ser obtidos a partir de 1 g de tecido, enquanto usando pronase, hialuronidase e colagenase - 4,3-10 6. Quando tratadas com colagenase, agrecan, proteínas, IL-6 e IL-8 permanecem na cultura de células em quantidades significativamente maiores do que com o tratamento sequencial com diferentes enzimas. Existem várias explicações para essas diferenças entre as duas culturas de células:
- Os receptores celulares são danificados ou inibidos por enzimas; o TGF-beta inibe a síntese de DNA e proteoglicanos em condrócitos recém-isolados (dia 1), enquanto a síntese de DNA e proteoglicanos em condrócitos cultivados em monocamada (7 dias) é estimulada pelo TGF-beta. No entanto, é necessário um período adequado para a reexpressão desses componentes da membrana antes do início do experimento.
- Proteases exógenas podem interromper a interação célula-matriz mediada por integrinas. A família das integrinas promove a ligação de condrócitos a moléculas da MEC (Shakibaei M. et al., 1997). Essa interrupção pode afetar a expressão de genes da matriz.
- Os restos de componentes da matriz podem regular a função sintética dos condrócitos. As integrinas são capazes de reconhecer os produtos da degradação da MEC, desempenhando assim um papel importante na reparação tecidual após a ação de enzimas proteolíticas. T. Larsson et al. (1989) relataram que a adição de proteoglicanos intactos ou fragmentados à cultura celular estimula a síntese de proteínas e proteoglicanos. No entanto, um alto nível de ácido hialurônico causa uma diminuição significativa na inclusão de sulfatos na síntese de proteoglicanos por condrócitos do embrião de galinha, condrócitos maduros do porco e células de condrossarcoma de rato. Além disso, o ácido hialurônico é um inibidor da liberação de proteoglicanos das células, mesmo na presença de IL-1b, TNF-a, FGF, o que indica a neutralização da primeira atividade biológica de fatores de crescimento e citocinas. O mecanismo exato subjacente à ação do ácido hialurônico permanece obscuro; Sabe-se que os condrócitos contêm um receptor para ácido hialurônico associado aos filamentos de actina do citosol. A ligação do ácido hialurônico ao seu receptor estimula a fosforilação de proteínas. Assim, esses dados demonstram a modulação da função metabólica dos condrócitos por moléculas fragmentadas ou nativas de proteínas da matriz por meio da ativação de receptores da membrana celular.
- A rápida estimulação da síntese de proteínas da matriz pelos condrócitos por enzimas pode ser consequência de alterações na forma dos condrócitos e/ou na reorganização do citoesqueleto.
- Algumas citocinas (p. ex., IL-8) e fatores de crescimento (p. ex., IGF-1, TGF-β) são sequestrados na MEC. O exemplo mais conhecido é a ligação do TGF-β pela decorina, que resulta na diminuição da capacidade deste último de induzir o crescimento celular em células ovarianas de hamster chinês. A descoberta de que o teor de decorina na cartilagem aumenta com a idade sugere uma diminuição da biodisponibilidade do TGF-β com o envelhecimento. Fatores de crescimento e citocinas podem ser liberados de resíduos da matriz durante a cultura e, subsequentemente, modular a função dos condrócitos.
Cultura de monocamada de condrócitos
O fenótipo diferenciado dos condrócitos é caracterizado principalmente pela síntese de colágeno tipo II e proteoglicanos específicos do tecido, bem como por um baixo nível de atividade mitótica. Há evidências de que, com o cultivo prolongado de células em monocamada, bem como após várias passagens celulares repetidas, os condrócitos perdem seus contornos esféricos e adquirem uma forma alongada, semelhante à de fibroblastos. Com essa metaplasia fibroblástica, a função sintética das células também é modificada, caracterizada por uma diminuição progressiva na síntese de colágenos tipos II, IX e XI e um aumento na síntese de colágenos tipos I, III e Y. Pequenos proteoglicanos não agregados são sintetizados devido ao agrecano funcional. A síntese de catepsina B e L é extremamente baixa em células diferenciadas, mas aumenta no processo de perda de diferenciação. A colagenase-1 é expressa em condrócitos diferenciados; com o cultivo prolongado, sua expressão diminui, enquanto a produção de inibidores teciduais de metaloproteases (TIMPs) aumenta.
Condrócitos diferenciados reexpressam colágeno do fenótipo diferenciado quando transferidos da monocamada para a cultura em suspensão. O processo de diferenciação provavelmente está relacionado ao formato da célula. Essa propriedade é regularmente utilizada por pesquisadores que estudam enxertos defeituosos com condrócitos autólogos. Um pequeno número de células obtidas a partir de material de biópsia pode ser expandido em cultura em monocamada e, em seguida, reintroduzido em uma matriz tridimensional antes do transplante. A reexpressão de um fenótipo específico por condrócitos desdiferenciados transferidos para cultura em agarose pode ser estimulada por TGF-β, complexo osseína-hidroxiapatita e ácido ascórbico.
Em resposta a fatores de crescimento e citocinas, os condrócitos são modificados durante o processo de diferenciação. A resposta celular a citocinas e fatores de crescimento difere entre condrócitos indiferenciados e diferenciados. A IL-1 estimula a proliferação de fibroblastos, enquanto o crescimento de condrócitos indiferenciados é inibido pela IL-1. A síntese de DNA é estimulada pelo IGF-1 em condrócitos alongados, mas não achatados. Em condrócitos diferenciados, os efeitos estimulatórios da IL-1β e do TNF-α na produção de procolagenase são mais pronunciados do que em condrócitos indiferenciados.
Cultivo de condrócitos
O cultivo de condrócitos em suspensão em meio líquido ou em matriz tridimensional natural ou sintética estabiliza o fenótipo dos condrócitos. As células mantêm sua forma esférica e sintetizam proteínas específicas do tecido. A cultura suspensa de condrócitos é geralmente recomendada para o estudo da formação de uma nova matriz pericelular. Culturas de condrócitos em polímeros absorventes sintéticos ou naturais são utilizadas para implantação de células em defeitos da cartilagem, a fim de estimular a regeneração do tecido cartilaginoso articular. O meio sintético ou natural para a implantação das células deve atender a uma série de requisitos:
- os implantes devem ter uma estrutura porosa para adesão e crescimento celular,
- nem o próprio polímero nem seus produtos de degradação devem causar inflamação ou reações tóxicas quando implantados in vivo,
- o portador do enxerto deve ter a capacidade de se ligar à cartilagem adjacente ou ao osso subcondral,
- a matriz natural ou sintética deve ter capacidade de absorção, sua degradação deve ser equilibrada pela regeneração tecidual,
- para facilitar o reparo da cartilagem, a estrutura química e a arquitetura dos poros da matriz devem facilitar a manutenção do fenótipo celular e a síntese de proteínas específicas do tecido pelos condrócitos nela colocados,
- Durante a implantação in vivo, é necessário estudar as propriedades mecânicas da matriz sintética ou natural.
 [ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ]
[ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ]
Suspensão de condrócitos em fase líquida
A adesão de células a vasos plásticos nos quais os condrócitos são cultivados pode ser evitada revestindo suas paredes com uma solução de metilcelulose, agarose, hidrogel (poli-2-hidroxietil metacrilato) ou uma mistura de colágeno-agarose. Nessas condições, os condrócitos formam aglomerados e sintetizam principalmente agrecano e colágenos específicos para tecidos (tipos II, IX e XI). Dois tipos de células são geralmente encontrados. As células localizadas no centro mantêm uma forma esférica e são circundadas por uma MEC bem desenvolvida, o que é confirmado por estudos histoquímicos e ultraestruturais. Na periferia, os condrócitos têm contornos discóides e são circundados por uma MEC esparsa; pouco se sabe sobre as características funcionais dessas células.
É possível cultivar condrócitos em microcarreadores mantidos em suspensão; esferas de dextrana (Cytodex), esferas de dextrana revestidas com colágeno (Cytodex III) e microesferas não porosas de colágeno tipo I (Cellagen) são utilizadas como microcarreadores. Nessas condições de cultivo, os condrócitos se fixam à superfície do microcarreador, mantêm sua forma esférica e produzem um material semelhante a uma matriz. Além disso, o uso de Cellagen promove a proliferação de condrócitos e a reexpressão do fenótipo normal. Portanto, o cultivo de condrócitos em microesferas de Cellagen pode ser utilizado para restaurar o fenótipo celular antes do transplante.
Outro método de cultivo de suspensões de condrócitos em meio líquido é o cultivo na forma de esferas densas constituídas por células (0,5-1 x 10 b ), obtidas por centrifugação. Esses condrócitos são capazes de produzir uma matriz contendo grande quantidade de proteoglicanos, colágeno tipo II, mas não colágeno tipo I, o que é confirmado por métodos histológicos, imuno-histoquímicos e quantitativos.
Suspensão de condrócitos na MEC natural
Os condrócitos podem ser cultivados em suspensão em uma matriz tridimensional (ágar mole, agarose, gel ou esponja de colágeno, ácido hialurônico, cola de fibrina, esferas de alginato).
Condrócitos cultivados em agarose mantêm seu fenótipo normal e sintetizam colágeno tipo II e agregados de agrecano específicos do tecido. Quando cultivados em agarose, os proteoglicanos sintetizados pela célula são liberados no meio por 50 dias. Para comparação, em uma cultura em monocamada, a fase celular é superpreenchida com glicosaminoglicanos já nos primeiros 5 a 6 dias de cultivo; quando cultivados no meio, após aumento da síntese e liberação de glicosaminoglicanos nos primeiros 8 a 10 dias, ocorre sua diminuição dependente do tempo. No entanto, o comportamento dos condrócitos quando cultivados em agarose difere daquele in vivo. Em agarose, um grande número de agregados de agrecano sintetizados contém moléculas menores e menos moléculas do que in vivo. O TGF-β estimula a síntese de proteoglicanos no explante, mas reduz a síntese de agrecano em agarose.
O alginato é um polissacarídeo linear obtido de algas marrons. Na presença de cátions divalentes, como íons Ca2 +, esse polímero se transforma em um gel. Cada condrócito aprisionado em alginato é circundado por uma matriz de polissacarídeos carregados negativamente, cujos poros são comparáveis aos da cartilagem hialina. A matriz formada pelos condrócitos em esferas de alginato consiste em dois compartimentos: uma fina camada de matriz associada à célula, correspondente à matriz pericelular e territorial da cartilagem articular, e uma matriz mais distante, equivalente à interterritorial do tecido nativo. No 30º dia de cultura, os volumes relativo e absoluto ocupados pelas células e por cada um dos dois compartimentos em uma esfera de alginato são quase completamente idênticos aos da cartilagem nativa. Por quase 30 dias, os condrócitos mantêm sua forma esférica e produzem agrecano, cujas propriedades hidrodinâmicas são semelhantes às das moléculas de agrecano na matriz da cartilagem articular, bem como às moléculas de colágeno dos tipos II, IX e XI. Ao mesmo tempo, como em outras culturas em suspensão, células achatadas estão presentes na superfície das esferas de alginato, que produzem uma pequena quantidade de moléculas de colágeno do tipo I, que são liberadas diretamente no meio e não são incorporadas à MEC. Observa-se proliferação moderada de condrócitos nas esferas de alginato. Após 8 meses de cultivo em gel de alginato, os condrócitos maduros não perdem a atividade metabólica e continuam a sintetizar colágeno tipo II e agrecano específicos para cada tecido.
H. Tanaka et al. (1984) investigaram as propriedades de difusão de várias moléculas naturais em alginato e descobriram que moléculas maiores que 70 kDa não se difundiram através do alginato. Portanto, a cultura de células em alginato é adequada para estudar a regulação da biossíntese da matriz e a organização da MEC. A disponibilidade de células cultivadas em alginato permite estudar a ação de fatores reguladores peptídicos e agentes farmacológicos nos níveis transcricional, pós-transcricional e translacional.
Os condrócitos também são cultivados em uma matriz de fibras de colágeno dos tipos I e II. S. Nehrer et al. (1997) compararam o funcionamento de condrócitos caninos em matrizes porosas de polímero de colágeno-proteoglicano contendo colágenos de diferentes tipos. Eles encontraram diferenças importantes na morfologia da função biossintética de condrócitos cultivados em matrizes de colágeno contendo colágeno tipos I e II. As células em uma matriz de colágeno tipo II mantiveram sua forma esférica, enquanto no colágeno tipo I elas tinham uma morfologia semelhante à de fibroblastos. Além disso, em uma matriz de colágeno tipo II, os condrócitos produziram uma quantidade maior de glicosaminoglicanos. J. van Susante et al. (1995) compararam as propriedades de condrócitos cultivados em gel de alginato e colágeno (tipo I). Os autores encontraram um aumento significativo no número de células no gel de colágeno, mas a partir do 6º dia de cultivo as células perderam seu fenótipo característico, transformando-se em células semelhantes a fibroblastos. No gel de alginato, observou-se uma diminuição no número de células, mas os condrócitos mantiveram seu fenótipo normal. No gel de colágeno, o número de proteoglicanos por célula foi significativamente maior do que no alginato, mas no gel observou-se uma diminuição na síntese de elementos da matriz a partir do 6º dia de cultivo, enquanto no alginato a síntese continuou a aumentar.
A matriz sólida tridimensional de fibrina é uma substância natural que sustenta os condrócitos suspensos nela, apresentando um fenótipo diferenciado. A matriz tridimensional de fibrina também pode ser usada como carreadora em transplantes de condrócitos. As vantagens da fibrina são a ausência de citotoxicidade, a capacidade de preencher espaços e a capacidade adesiva. Estudos histológicos e bioquímicos, autorradiografia e microscopia eletrônica demonstraram que os condrócitos em gel de fibrina mantêm sua morfologia, proliferam e produzem matriz mesmo após 2 semanas de cultivo. No entanto, G. Homminga et al. (1993) relataram que a desintegração da fibrina começa após 3 dias de cultivo, e a desdiferenciação dos condrócitos progride.
Suspensão de condrócitos em ECM artificial (sintética)
Implantes de cartilagem para cirurgia reconstrutiva ou ortopédica podem ser obtidos pelo cultivo de condrócitos isolados in vitro em uma matriz sintética biocompatível.
Condrócitos cultivados em ácido poliglicólico proliferam e mantêm morfologia e fenótipo normais por 8 semanas. O complexo condrócito-ácido poliglicólico consiste em células, glicosaminoglicanos, colágenos e possui uma cápsula externa de colágeno. No entanto, esses implantes contêm dois tipos de moléculas de colágeno: I e II. Implantes de condrócitos desdiferenciados por uma série de passagens apresentam maior quantidade de glicosaminoglicanos e colágenos do que implantes de condrócitos primariamente indiferenciados.
L. Freed et al. (1993b) compararam o comportamento de culturas de condrócitos humanos e bovinos em ácido poliglicólico fibroso (FPGA) e ácido polilático poroso (PPLA). Após 6 a 8 semanas de cultivo de condrócitos bovinos em FPGA ou PPLA, os autores observaram proliferação celular e regeneração da matriz cartilaginosa. Em FPGA, os condrócitos tinham formato esférico e estavam localizados em lacunas circundadas por uma matriz cartilaginosa. Após 8 semanas de cultivo in vitro, o tecido regenerado continha até 50% de matéria seca (4% de massa celular, 15% de glicosaminoglicanos e 31% de colágenos). Em PPLA, as células tinham formato fusiforme e uma pequena quantidade de glicosaminoglicanos e colágeno. Em FPGA, o crescimento celular foi 2 vezes mais intenso do que em PPLA. In vivo, condrócitos cultivados em VPGK e PPLC produziram tecido histologicamente semelhante à cartilagem em 1 a 6 meses. Os implantes continham glicosaminoglicanos, colágenos tipo I e II.
Condrócitos fetais bovinos foram cultivados em polietileno poroso de alta densidade, hidrofóbico e hidrofílico. Após 7 dias de incubação em ambos os substratos, as células mantiveram o formato esférico e continham principalmente colágeno tipo II. Após 21 dias de cultivo, constatou-se que a matriz hidrofílica continha mais colágeno tipo II do que a matriz hidrofóbica.
O tecido cartilaginoso também pode ser obtido por cultivo em monocamada sobre filtros Millicell-CM. O pré-revestimento dos filtros com colágeno é necessário para a fixação das condroitinas. O exame histológico da cultura demonstra o acúmulo de condrócitos na MEC contendo proteoglicanos e colágeno tipo II. O colágeno tipo I não foi detectado em tal cultura. Os condrócitos no tecido cartilaginoso obtido têm formato esférico, mas na superfície do tecido são um tanto achatados. A espessura do tecido recém-formado aumentou com o tempo e dependeu da densidade inicial da monocamada de células. Em condições ótimas de cultivo, a espessura do tecido cartilaginoso atingiu 110 μm, e a organização de suas células e colágeno em camadas superficiais e profundas é semelhante à da cartilagem articular. A MEC contém aproximadamente 3 vezes mais colágeno e proteoglicanos. Após 2 semanas de cultivo, observou-se acúmulo de matriz, permitindo que o tecido fosse extraído do filtro e utilizado para transplante.
Sims et al. (1996) estudaram o cultivo de condrócitos em gel de óxido de polietileno, uma matriz polimérica encapsulada que permite a transferência de um grande número de células por injeção. Seis semanas após a injeção no tecido subcutâneo de camundongos atímicos, formou-se nova cartilagem, caracterizada morfologicamente por uma opalescência branca semelhante à cartilagem hialina. Dados histológicos e bioquímicos indicaram a presença de condrócitos em proliferação ativa, produzindo MEC.
Explicação
A explantação de tecido cartilaginoso é usada para estudar os processos de anabolismo e catabolismo, manutenção da homeostase, reabsorção e reparo. Os condrócitos em explantes de cartilagem mantêm um fenótipo normal e composição de MEC semelhantes aos da cartilagem articular in vivo. Após 5 dias de cultivo na presença de soro, um nível constante de processos de síntese e degradação natural é alcançado. A reabsorção tecidual pode ser acelerada na cultura principal e em cultura com a adição de soro usando vários agentes, como IL-IB, TNF-a, lipopolissacarídeos bacterianos, derivados do ácido retinoico ou radicais ativos de oxigênio. Para estudar a reparação da cartilagem, seu dano é induzido por mediadores inflamatórios solúveis (H2O2, IL 1, TNF-a) ou ruptura física da matriz.
O método de cultura organotípica é um modelo para estudos in vitro dos efeitos de fatores externos isolados sobre os condrócitos e a matriz circundante. In vivo, os condrócitos estão localizados esparsamente na MEC e não entram em contato uns com os outros. A cultura do explante de cartilagem articular preserva essa organização estrutural, bem como as interações específicas entre os condrócitos e o ambiente extracelular circundante. Este modelo também é utilizado para estudar os efeitos do estresse mecânico, agentes farmacológicos, fatores de crescimento, citocinas e hormônios no metabolismo da cartilagem.
Outra vantagem da remoção do tecido cartilaginoso é a ausência de danos aos condrócitos sob a ação de enzimas proteolíticas ou fatores mecânicos, o que é inevitável ao isolar células. Receptores e outras proteínas e glicoproteínas de membrana são protegidos de fatores danosos.
[ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ]
Cultura de condrões
O côndron é uma unidade estrutural, funcional e metabólica da cartilagem articular, constituída por um condrócito, sua matriz pericelular e cápsula filamentosa compacta, sendo responsável pela homeostase da matriz. Os côndrons são extraídos mecanicamente da cartilagem e coletados por meio de várias homogeneizações sucessivas de baixa velocidade. Os côndrons isolados de zonas com diferentes profundidades de cartilagem podem ser divididos em quatro categorias: côndrons simples, côndrons pareados, côndrons múltiplos (três ou mais) dispostos linearmente (colunas de côndrons) e côndrons agrupados.
Condrões únicos são geralmente encontrados nas camadas médias da cartilagem intacta, condrões pareados são encontrados na borda das camadas média e profunda, condrões múltiplos dispostos linearmente são típicos das camadas profundas da cartilagem intacta. Finalmente, os aglomerados de condrões consistem em grupos organizados aleatoriamente de condrões únicos e pareados, que retêm um estado agregado após a homogeneização. Os aglomerados de condrões são grandes fragmentos de cartilagem, geralmente contendo vários condrões e fibrilas de colágeno dispostas radialmente, ou seja, uma organização típica característica das camadas profundas da matriz. Os condrões são imobilizados em agarose transparente, o que permite estudos de sua estrutura, composição molecular e atividade metabólica. O sistema condrão-agarose é considerado um micromodelo de cartilagem, que difere do sistema condrócito-agarose tradicional por preservar o microambiente natural e não haver necessidade de sintetizá-lo e montá-lo. A cultura de condrões é um modelo para estudar as interações de células e matriz na cartilagem articular em condições normais e patológicas.
[ 22 ], [ 23 ], [ 24 ], [ 25 ], [ 26 ], [ 27 ]
Cultura de condrócitos imortais
Vírus contendo DNA recombinante ou oncogenes capazes de tornar uma célula "imortal" são utilizados para criar linhagens celulares permanentes. Condrócitos imortais têm a capacidade de proliferar indefinidamente, mantendo um fenótipo estável. F. Mallein-Gerin et al. (1995) demonstraram que o oncogene SV40T induz a proliferação de condrócitos de camundongos, que continuam a expressar de forma estável os colágenos tipos II, IX e XI, bem como agrecano articular e proteína de ligação. No entanto, essa linhagem celular adquire a capacidade de sintetizar colágeno tipo I ao ser cultivada em monocamada ou em gel de agarose.
W. Horton et al. (1988) descreveram uma linhagem de células imortais com baixo nível de expressão de mRNA de colágeno tipo II. Essas células foram obtidas por transformação com um retrovírus de camundongo contendo os oncogenes I-myc e y-ra. Esse tipo de célula representa um modelo único para o estudo das interações da matriz articular na ausência de colágeno tipo II, bem como da regulação da síntese de colágeno tipo II.
A cultura de condropritos com genes mutados ou deletados é um modelo conveniente para o estudo de sua função fisiológica. Este modelo é especialmente adequado para estudar o papel de moléculas específicas na organização da matriz da cartilagem ou para investigar os efeitos de vários fatores regulatórios no metabolismo da cartilagem. Condrócitos com um gene deletado para colágeno tipo IX sintetizam fibrilas de colágeno que são mais largas do que o normal, indicando que o colágeno tipo IX regula o diâmetro das fibrilas. Conforme observado no Capítulo 1, uma mutação no gene COLAI que codifica o colágeno tipo II foi recentemente descoberta em famílias com osteoartrite generalizada primária. Para estudar o efeito do colágeno tipo II mutante na matriz articular, R. Dharmrvaram et al. (1997) transfectaram (infectaram com ácido nucleico estranho) COL 2 AI defeituoso (arginina na posição 519 é substituída por cisteína) em condrócitos fetais humanos in vitro.
Sistema de cocultura. Na articulação, a cartilagem interage com outros tipos de células presentes na membrana sinovial, no líquido sinovial, nos ligamentos e no osso subcondral. O metabolismo dos condrócitos pode ser afetado por diversos fatores solúveis sintetizados por essas células. Assim, na artrite, a cartilagem articular é destruída por enzimas proteolíticas e radicais livres produzidos pelas células sinoviais. Por isso, modelos foram desenvolvidos para estudar interações complexas entre a cartilagem e os tecidos circundantes, denominadas coculturas.
S. Lacombe-Gleise et al. (1995) cultivaram condrócitos e osteoblastos de coelho em um sistema de cocultura (COSTAR), no qual as células foram separadas por uma membrana microporosa (0,4 μm), permitindo a troca entre os dois tipos celulares sem qualquer contato direto. Este estudo demonstrou a capacidade dos osteoblastos de estimular o crescimento de condrócitos por meio de mediadores solúveis.
AM Malfait e coautores (1994) estudaram a relação entre monócitos e condrócitos do sangue periférico. Este modelo é conveniente para o estudo de processos mediados por citocinas em artropatias inflamatórias (artrite reumatoide, espondiloartrites soronegativas, etc.). Os autores do modelo separaram as células com uma membrana de ligação a proteínas com poros de 0,4 μm de diâmetro. O estudo mostrou que monócitos estimulados com lipopolissacarídeo produziram IL-1 e TNF-α, que inibiram a síntese de agrecano pelos condrócitos e contribuíram para a degradação de agregados de agrecano já sintetizados.
K. Tada et al. (1994) criaram um modelo de cocultura no qual células endoteliais em um gel de colágeno (tipo I) foram colocadas em uma câmara interna separada da câmara externa, com condrócitos colocados nela por um filtro com poro de 0,4 μm. Em um estado de isolamento completo da câmara externa, células endoteliais humanas formaram tubos em um gel de colágeno na presença de EGF ou TGF-α. Quando ambos os tipos de células foram cultivados simultaneamente, a formação de tubos dependentes de TGF-α pelas células endoteliais foi inibida. A inibição desse processo pelos condrócitos foi parcialmente eliminada por anticorpos anti-TGF-β. Pode-se presumir que o TGF-β produzido pelos condrócitos inibe a vascularização da própria cartilagem.
S. Groot et al. (1994) cultivaram simultaneamente condrócitos das zonas hipertrófica e proliferativa do osso de um feto de camundongo de 16 dias de idade com pedaços de tecido cerebral. Após 4 dias de cultivo, observou-se a transdiferenciação de condrócitos em osteoblastos e o início da formação de osteoides. Após 11 dias de cultivo, parte da cartilagem foi substituída por tecido ósseo e a matriz óssea foi parcialmente calcificada. Alguns neuropeptídeos e neurotransmissores produzidos pelo tecido cerebral afetam o metabolismo dos osteoblastos ou possuem receptores para eles. Entre eles estão a norepinefrina, o peptídeo intestinal vasoativo, o peptídeo relacionado ao gene da calcitonina, a substância P e a somatostatina. Pedaços de tecido cerebral cultivados juntamente com condrócitos podem produzir alguns dos fatores listados capazes de induzir o processo de transdiferenciação de condrócitos em osteoblastos.
[ 28 ], [ 29 ], [ 30 ], [ 31 ], [ 32 ], [ 33 ]
A influência de fatores externos na cultura de condrócitos
A influência da tensão de oxigênio no metabolismo dos condrócitos
Na maioria dos casos, as culturas de condrócitos se desenvolvem sob condições de tensão atmosférica de oxigênio. No entanto, sabe-se que os condrócitos in vivo existem em condições hipóxicas e a tensão de oxigênio varia sob várias condições patológicas. Durante o processo de maturação, são observadas alterações significativas no suprimento sanguíneo para as epífises. Como a vascularização varia em diferentes zonas da placa de crescimento, a tensão de oxigênio nelas também varia. C. Brighton e R. Heppenstall (1971) demonstraram que, na placa tibial de coelhos, a tensão de oxigênio na zona hipertrófica é menor do que na cartilagem circundante. Medições de alguns parâmetros metabólicos mostraram que os condrócitos são capazes de responder rapidamente a mudanças locais na concentração de oxigênio. Primeiramente, em baixa tensão de oxigênio, seu consumo pelos condrócitos diminui. Com uma diminuição na tensão de oxigênio de 21% para 0,04%, a utilização de glicose aumenta, a atividade de enzimas glicolíticas e a síntese de ácido lático aumentam. Mesmo em baixa tensão de oxigênio, a quantidade absoluta de ATP, ADP e AMP permanece estável. Esses dados indicam que o metabolismo dos condrócitos visa a conservação máxima de energia. No entanto, a atividade sintética e, portanto, os processos de reparação, alteram-se em condições hipóxicas.
A alta tensão de oxigênio também afeta o metabolismo dos condrócitos, causando diminuição da síntese de proteoglicanos e DNA e degradação da matriz da cartilagem. Esses efeitos geralmente são acompanhados pela produção de radicais livres de oxigênio.
A influência da concentração de íons e da pressão osmótica do ambiente na função dos condrócitos
Na cartilagem nativa, a concentração de íons difere significativamente daquela em outros tecidos: o teor de sódio no meio extracelular é de 250-350 mmol e sua osmolaridade é de 350-450 mosmol. Quando os condrócitos são isolados da MEC e incubados em meio padrão (DMEM (Meio Essencial Mínimo de Dulbecco), com osmolaridade de 250-280,7 mosmol), o ambiente ao redor das células muda drasticamente. Além disso, a concentração de cálcio e potássio no meio padrão é significativamente menor do que no tecido nativo, e a concentração de ânions é significativamente maior.
A adição de sacarose ao meio aumenta sua osmolaridade e induz um aumento intracelular transitório na concentração de ânions H + e cálcio no citosol. Tais alterações intracelulares podem afetar os processos de diferenciação dos condrócitos e sua atividade metabólica. J. Urban et al. (1993) descobriram que a incorporação de 35 8-sulfato e 3 H-prolina por condrócitos isolados incubados em DMEM padrão por 2-4 h foi de apenas 10% daquela no tecido nativo. A intensidade da síntese atingiu um máximo em uma osmolaridade do meio extracelular de 350-400 mosmol tanto em condrócitos recém-isolados quanto em explantes de tecido cartilaginoso. Além disso, o volume de condrócitos aumentou em 30-40% após a colocação das células isoladas em DMEM padrão com a osmolaridade especificada. Entretanto, ao cultivar condrócitos em condições de osmolaridade não fisiológica por 12 a 16 horas, as células se adaptam às novas condições, reduzindo a intensidade da biossíntese proporcionalmente à mudança na osmolaridade do ambiente extracelular.
P. Borgetti et al. (1995) estudaram o efeito da osmolaridade do meio extracelular no crescimento, morfologia e biossíntese de condrócitos suínos. Os autores demonstraram características bioquímicas e morfológicas semelhantes em condrócitos cultivados em meios com osmolaridade de 0,28 e 0,38 mosmol. Em uma osmolaridade média de 0,48 mosmol, foi observada uma diminuição na proliferação celular e na síntese proteica durante as primeiras 4 a 6 horas de cultivo, mas esses parâmetros se recuperaram posteriormente e, eventualmente, atingiram os valores de controle. Quando os condrócitos foram cultivados em um meio com osmolaridade de 0,58 mosmol, as células perderam a capacidade de manter a intensidade fisiológica dos processos proliferativos e, após 6 dias, o número de condrócitos foi significativamente reduzido. Em uma osmolaridade média de 0,58 mosmol, foi observada profunda inibição da síntese proteica. Além disso, quando cultivados em meios com osmolaridade de 0,28-0,38 mOsm, os condrócitos mantêm seu fenótipo fisiológico; em osmolaridade mais alta (0,48-0,58 mOsm), ocorrem alterações significativas na morfologia celular, manifestadas pela perda do fenótipo característico, pela transformação dos condrócitos em células semelhantes a fibroblastos e pela perda da capacidade das células de montar proteoglicanos da matriz. Os resultados deste estudo indicam a capacidade dos condrócitos de responder a flutuações limitadas na osmolaridade do ambiente extracelular.
Mudanças na concentração de outros íons também podem afetar os processos de biossíntese em condrócitos. Assim, o grau de incorporação de 35 S (sulfato) aumenta pela metade com um aumento na concentração de íons potássio de 5 mmol (a concentração no meio DM EM padrão) para 10 mmol (a concentração na MEC in vivo). Concentrações de cálcio abaixo de 0,5 mmol promoveram a produção de colágeno por condrócitos bovinos maduros, enquanto uma concentração de 1-2 mmol (correspondente à concentração no meio DM EM padrão) causou uma diminuição significativa na síntese de colágeno. Um aumento moderado na biossíntese foi observado em altos níveis de cálcio (2-10 mmol). Vários cátions participam da ligação dos condrócitos às proteínas da MEC. Assim, os íons magnésio e manganês fornecem ligação à fibronectina e ao colágeno tipo II, enquanto os íons cálcio não participam da ligação dos condrócitos às proteínas. Assim, os resultados dos estudos descritos indicam a influência das alterações nos íons extracelulares de potássio, sódio, cálcio e osmolaridade do meio na função biossintética de condrócitos incubados em meios padrão.
Efeito do estresse mecânico no metabolismo dos condrócitos
A imobilização articular causa atrofia reversível da cartilagem, o que indica a necessidade de estímulos mecânicos para os processos metabólicos normais na MEC. Na maioria dos casos, os modelos de cultura de células utilizados existem sob pressão atmosférica normal. M. Wright et al. (1996) mostraram que o ambiente mecânico afeta o metabolismo dos condrócitos; a resposta celular depende da intensidade e frequência da carga compressiva. Experimentos com carga em explantes de cartilagem articular intacta in vitro demonstraram uma diminuição na síntese de proteínas e proteoglicanos sob a ação de carga estática, enquanto a carga dinâmica estimula esses processos. Os mecanismos exatos do efeito da carga mecânica na cartilagem são complexos e provavelmente estão associados à deformação celular, pressão hidrostática, pressão osmótica, potencial elétrico e receptores celulares de superfície para moléculas da matriz. Para estudar o efeito de cada um desses parâmetros, é necessário criar um sistema no qual um parâmetro possa ser variado independentemente. Por exemplo, a cultura de explantes não é adequada para estudar a deformação celular, mas pode ser usada para estudar o efeito geral da pressão na atividade metabólica dos condrócitos. A compressão da cartilagem leva à deformação celular e também é acompanhada pelo surgimento de um gradiente de pressão hidrostática, potencial elétrico, fluxo de fluido e alterações em parâmetros físico-químicos como o teor de água na matriz, densidade de carga elétrica e nível de pressão osmótica. A deformação celular pode ser estudada utilizando condrócitos isolados imersos em gel de agarose ou colágeno.
Vários sistemas foram desenvolvidos para estudar o efeito da estimulação mecânica na cultura de condrócitos. Alguns pesquisadores usam sistemas nos quais a pressão é aplicada à cultura de células através da fase gasosa. Assim, JP Veldhuijzen et al. (1979), usando pressão de 13 kPa acima da atmosférica com baixa frequência (0,3 Hz) por 15 min, observaram um aumento na síntese de AMPc e proteoglicanos e uma diminuição na síntese de DNA. R. Smith et al. (1996) mostraram que a exposição intermitente de uma cultura de condrócitos bovinos primários à pressão hidrostática (10 MPa) com frequência de 1 Hz por 4 h causou um aumento na síntese de agrecano e colágeno tipo II, enquanto a pressão constante não afetou esses processos. Usando um sistema semelhante, Wright et al. (1996) relataram que a pressão cíclica na cultura de células está associada à hiperpolarização da membrana celular dos condrócitos e à ativação de canais de potássio dependentes de Ca 2+. Assim, os efeitos da pressão cíclica são mediados por canais iônicos ativados por estiramento na membrana dos condrócitos. A resposta dos condrócitos à pressão hidrostática depende das condições de cultura celular e da frequência da carga aplicada. Assim, a pressão hidrostática cíclica (5 MPa) diminui a incorporação de sulfato na monocamada de condrócitos nas frequências de 0,05, 0,25 e 0,5 Hz, enquanto em frequências superiores a 0,5 Hz, a incorporação de sulfato no explante de cartilagem aumenta.
M. Bushmann et al. (1992) relataram que condrócitos em géis de agarose alteram a biossíntese em resposta à carga mecânica estática e dinâmica da mesma forma que o órgão intacto cultivado. Os autores descobriram que a carga mecânica gera um estímulo hiperosmótico com subsequente diminuição do pH nos condrócitos.
O efeito do estiramento mecânico pode ser estudado em uma cultura de células imersa em um gel. A força de estiramento pode ser criada usando um vácuo controlado por computador. Quando o sistema está sob um certo grau de vácuo, o fundo da placa de Petri com a cultura de células é estendido por uma quantidade conhecida, a deformação é máxima nas bordas do fundo da placa e mínima no centro. O estiramento também é transmitido aos condrócitos cultivados na placa de Petri. Usando este método, K. Holm-vall et al. (1995) mostraram que em células de condrossarcoma cultivadas em um gel de colágeno (tipo II), a expressão de mRNA de uma integrina 2 β é aumentada . Umaintegrina 2 β é capaz de se ligar ao colágeno tipo II. É considerado um mecanorreceptor, uma vez que interage com proteínas de ligação à actina, conectando assim a MEC e o citoesqueleto.
O efeito do pH no metabolismo dos condrócitos
O pH do fluido intersticial da MEC do tecido cartilaginoso é mais ácido do que em outros tecidos. A. Maroudas (1980) determinou o pH da matriz da cartilagem articular em 6,9. B. Diamant et al. (1966) encontraram um pH de 5,5 em condições patológicas. Sabe-se que os condrócitos vivem em baixa PO₂, o que indica o importante papel da glicólise (95% de todo o metabolismo da glicose) no metabolismo dessas células; a glicólise é acompanhada pela produção de grandes quantidades de ácido lático.
Além da acidificação do ambiente pelos produtos da glicólise, os próprios componentes da matriz são de grande importância. A grande quantidade de carga negativa fixa nos proteoglicanos modifica a composição iônica extracelular: observa-se uma alta concentração de cátions livres (por exemplo, H +, Na +, K + ) e uma baixa concentração de ânions (por exemplo, O2, HCO3 ). Além disso, sob a influência da carga mecânica, a água é expelida da MEC, o que leva a um aumento na concentração de cargas negativas fixas e à atração de mais cátions para a matriz. Isso é acompanhado por uma diminuição do pH do ambiente extracelular, que afeta o pH intracelular, modificando assim o metabolismo dos condrócitos. R. Wilkin e A. Hall (1995) estudaram o efeito do pH do ambiente extracelular e intracelular na biossíntese da matriz por condrócitos bovinos isolados. Eles observaram uma modificação dupla da síntese da matriz com a diminuição do pH. Uma ligeira redução no pH (7,4
[ 34 ], [ 35 ], [ 36 ], [ 37 ], [ 38 ], [ 39 ], [ 40 ]
Influência da composição do meio de cultura no metabolismo dos condrócitos
O meio de cultura de condrócitos deve corresponder às condições experimentais. Nos últimos anos, o soro bovino tem sido utilizado para otimizar as condições de cultura. No entanto, ao utilizar soro, alguns pontos importantes devem ser considerados:
- crescimento externo de células da periferia do tecido em culturas de órgãos,
- variabilidade na composição de soros de diferentes séries,
- a presença de componentes desconhecidos neles,
- aumento do risco de interferências e artefatos ao estudar a influência de vários fatores biológicos na atividade metabólica das células.
Um exemplo deste último é um estudo do efeito do EGF em condrócitos da cartilagem em ratos. O EGF estimulou a incorporação de 3H -timidina e um aumento no conteúdo de DNA na cultura. Este efeito foi mais pronunciado em baixas concentrações séricas (<1%), mas em altas concentrações (>7,5%) o efeito desapareceu.
É bem conhecido que os níveis de síntese e degradação em DMEM suplementado com soro de bezerro são significativamente aumentados em comparação com as condições in vivo. As diferenças entre o metabolismo in vivo e in vitro podem ser devidas a diferenças entre o fluido sinovial e o meio em que as células são cultivadas. Lee et al. (1997) cultivaram condrócitos bovinos jovens em agarose usando um meio nutriente contendo DMEM suplementado com 20% de soro de bezerro e uma grande quantidade de fluido sinovial alogênico normal. A presença de fluido sinovial no meio induziu um aumento na quantidade de proteoglicanos, até 80% da quantidade total de fluido sinovial. Esses resultados indicam que o fluido sinovial em cultura induz um nível de metabolismo semelhante ao in vivo, com um alto nível de síntese de glicosaminoglicanos e um baixo nível de divisão celular.
G. Verbruggen et al. (1995) demonstraram que a síntese de 35 S-arrpeKaHa por condrócitos humanos cultivados em agarose em meio DMEM isento de soro foi de 20 a 30% do nível de síntese observado em meio DMEM suplementado com 10% de soro bovino. Os autores determinaram a extensão em que IGF-1, IGF-2, TGF-R ou insulina restauraram a produção de agrecano em meio isento de soro. Os autores concluíram que 100 ng/ml de insulina, IGF-1 ou IGF-2 restauraram parcialmente a síntese de agrecano para 39 a 53% do nível de controle. Nenhum sinergismo ou acumulação foi observado com a combinação dos fatores listados. Ao mesmo tempo, 10 ng/ml de TGF-R na presença de 100 ng/ml de insulina estimularam a síntese de agrecano para 90% ou mais do nível de referência. Finalmente, a transferrina sérica humana, isoladamente ou em combinação com insulina, não afetou a síntese de agrecano. Quando o soro bovino foi substituído por albumina sérica bovina, o conteúdo de agregados de agrecano diminuiu significativamente. O enriquecimento do meio de cultura com insulina, IGF ou TGF-R restaurou parcialmente a capacidade das células de produzir agregados de agrecano. Além disso, o IGF-1 e a insulina são capazes de manter a homeostase em culturas celulares. Após 40 dias de cultivo em meio enriquecido com 10-20 ng/ml de IGF-1, a síntese de proteoglicanos foi mantida no mesmo nível ou até mais alta em comparação ao meio contendo 20% de soro bovino. Os processos catabólicos ocorreram mais lentamente no meio enriquecido com IGF-1 do que no meio enriquecido com solução de albumina a 0,1%, mas um pouco mais rápido no meio enriquecido com 20% de soro. Em culturas de longa duração, 20 ng/ml de IGF-1 mantêm um estado estável das células.
D. Lee et al. (1993) compararam o efeito da composição do meio de cultura (DMEM, DMEM + 20% de soro bovino, DMEM + 20 ng/ml de IGF-1) na síntese de DNA em uma cultura de explante de tecido cartilaginoso, uma cultura em monocamada e em suspensão de agarose. Ao cultivar em agarose na presença de soro, os autores observaram uma tendência dos condrócitos se agruparem em grandes aglomerados. As células cultivadas sem soro ou com IGF-1 mantiveram uma forma arredondada em agarose, coletadas em pequenos grupos, mas não formaram grandes agregados. Em uma monocamada, a síntese de DNA foi significativamente maior no meio contendo soro do que no meio enriquecido com IGF-1; a síntese de DNA neste último foi significativamente maior do que no meio não enriquecido. Não foram encontradas diferenças na síntese de DNA quando os condrócitos foram cultivados em suspensão de agarose em um meio não enriquecido e em um meio com IGF-1. Ao mesmo tempo, o cultivo de suspensões de condrócitos em agarose em meio enriquecido com soro foi acompanhado por maior incorporação do radionucleotídeo 3 H-timidina em comparação a outros meios.
A vitamina C é necessária para a ativação de enzimas envolvidas na formação de uma estrutura helicoidal estável das fibrilas de colágeno. Condrócitos deficientes em ácido ascórbico sintetizam precursores de colágeno não helicoidal sub-hidroxilados, que são secretados lentamente. A administração de ácido ascórbico (50 μg/ml) causa hidroxilação dos colágenos tipos II e IX e sua secreção em quantidades normais. A adição de vitamina C não afetou o nível de síntese de proteoglicanos. Portanto, a secreção de colágeno é regulada independentemente da secreção de proteoglicanos.